Содержание
Морфологические и физиологические особенности плода в разные периоды внутриутробного развития
Похожие презентации:
Внутриутробное развитие плода
Внутриутробное развитие плода
Внутриутробное развитие плода: эмбриональная и фетальная фаза развития плода, «критические периоды» развития
Этапы развития внутриутробного плода и физиологические изменения в различные месяцы беременности
Анатомо-физиологические особенности детей различных возрастных периодов
Внутриутробное развитие плода (8-16 неделя)
Период беременности и родов. Оплодотворение
Критические периоды в ходе онтогенеза человека. Пагубное влияние на развитие плода лекарственных препаратов, алкоголя, никотина
Оплодотворение. Этапы внутриутробного развития влияние патогенных факторов на эмбрион и плод
Анатомо-физиологические особенности органов и систем в различные периоды детства
1. Морфологические и физиологические особенности плода в разные периоды внутриутробного развития
МЛ-502ЛУЦЕНКО А.
 М.
М.2. 1 – 4 неделя
1 – 4 неделяДробление зиготы
Имплантация
Образование зародыша
Образование зачатков важнейших органов и
зародышевых оболочек
3. 7 – 8 недель
Длина плода 3-3,5 смТело сформировано
Имеются зачатки конечностей
Головка равняется длине туловища, не ней заметны
зачатки глаз, носа, рта
Начинается формирование половых органов
Регистрируется сердцебиение плода
4. 11 – 12 недель
Длина плода 8-9 см, масса 20-25 г.Головка крупная
Заметно различие в строении наружных половых
органов
Конечности совершают движения
Видны пальцы рук и ног
В хрящевом зачатке скелета появляются первые точки
окостенения
5. 15 – 16 недель
Длина плода 16 см, масса 110-120 г.Формируется лицо
Происходит окостенение черепа
В основном заканчивается формирование
мышечной системы
Движения конечностей становятся активнее, но
матерью не воспринимаются
Пол плода различается ясно
6.
 19 – 20 недельПлод достигает длины 25-26 см, масса 280-300 г.
19 – 20 недельПлод достигает длины 25-26 см, масса 280-300 г.Кожа красная, покрывается пушковыми волосами
Сальные железы начинают выделять жировое
вещество
С 18-20 недели движения плода ощущаются матерью
При аускультации можно прослушать сердцебиение
плода
7. 23 – 24 недели
Длина плода 30 см, масса 600-680 г.Движения энергичные
Плод может родиться живым
Может делать дыхательные движения
Плод, родившийся после 22 недели массой более 500 г.
— подлежит выхаживания и регистрации как
живорожденный.
8. 27 – 28 недель
Длина плода 35 см, масса 1000-1200 г.Подкожный жир развит слабо
Кожа морщинистая
На всем теле пушковидные волосы
Ушные и носовые хрящи мягкие, ногти не доходят до концов
пальцев рук и ног
У мальчиков: яички не опустились в мошонку; у девочек: малые
половые губы не прикрыты большими
Плод рождается живым, дышит, но мало жизнеспособен.
Нуждается в интенсивном уходе.
9.
 31 – 32 неделяДлина плода 40-42 см, масса 1500-1700 г.
31 – 32 неделяДлина плода 40-42 см, масса 1500-1700 г.Плод рождается жизнеспособный но требует
особого ухода
10. 35 – 36 недель
Длина плода 45-48 см, масса 2400-2500 г.ПЖК слой увеличивается
Кожа гладкая, розовая
Пушковых волос на теле меньше
Плод, родившийся в этот срок, жизнеспособен, громко
кричит, открывает глаза, выражен сосательный
рефлекс.
English Русский Правила
29. Морфологические и физиологические особенности плода в разные периоды внутриутробной жизни. Понятие и критерии жизнеспособного плода.
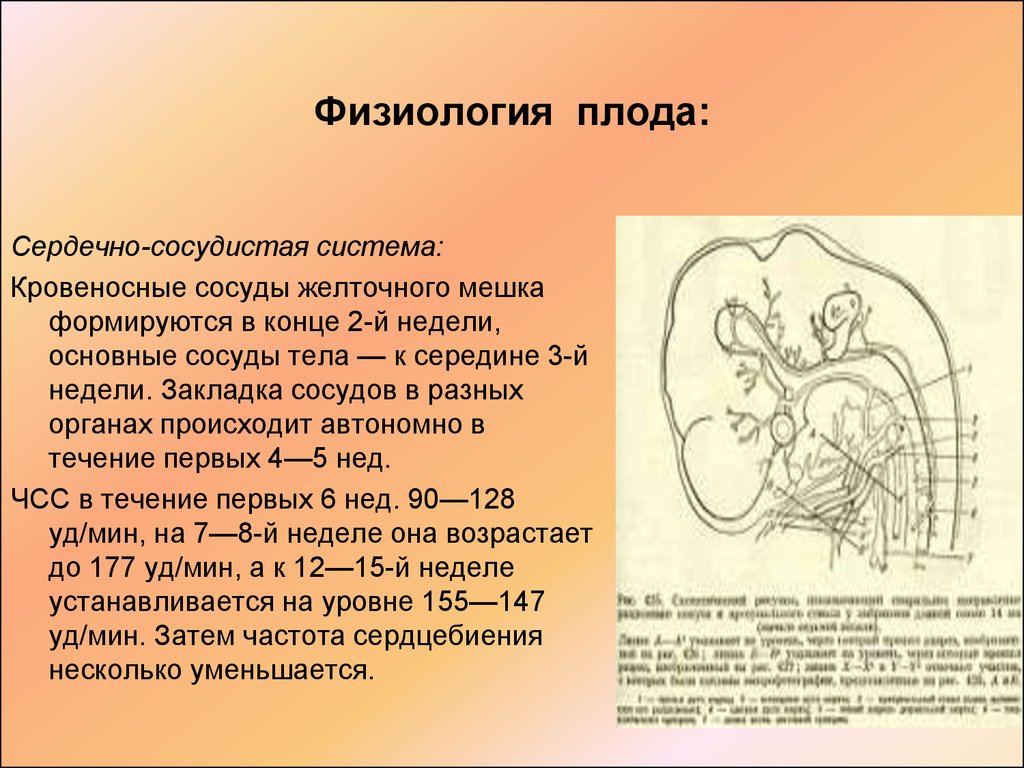
Средняя продолжительность беременности составляет 280 дней (40 нед, или 10 акушерских месяцев).
В
течение I месяца внутриутробного развития
совершаются процесс дробления зиготы,
возникают морула, бластула и бластоцист.
На стадии бластоцисты -имплантация
зародыша в децидуальную оболочку матки,
далее закладка и дифференцировка органов
и систем эмбриона, а также образование
зародышевых оболочек.
плодное яйцо может быть визуализировано со срока беременности 2-3 нед-диаметр плодного яйца =2-4 мм.
Сердце бьется с 21 дня
На 7-й неделе длина эмбриона = 5-9 мм, к концу 7-й недели возможна визуализациянервной трубки, закладка ссс, ЦНС, половой системы
К концу 8-й недели -определяются конечности и ЧСС-112-136 уд/мин. Четкая идентификация эмбриона (головка, туловище, конечности) с 8-9 нед.
12 нед- масса 20-25гр длина — 6-7 см
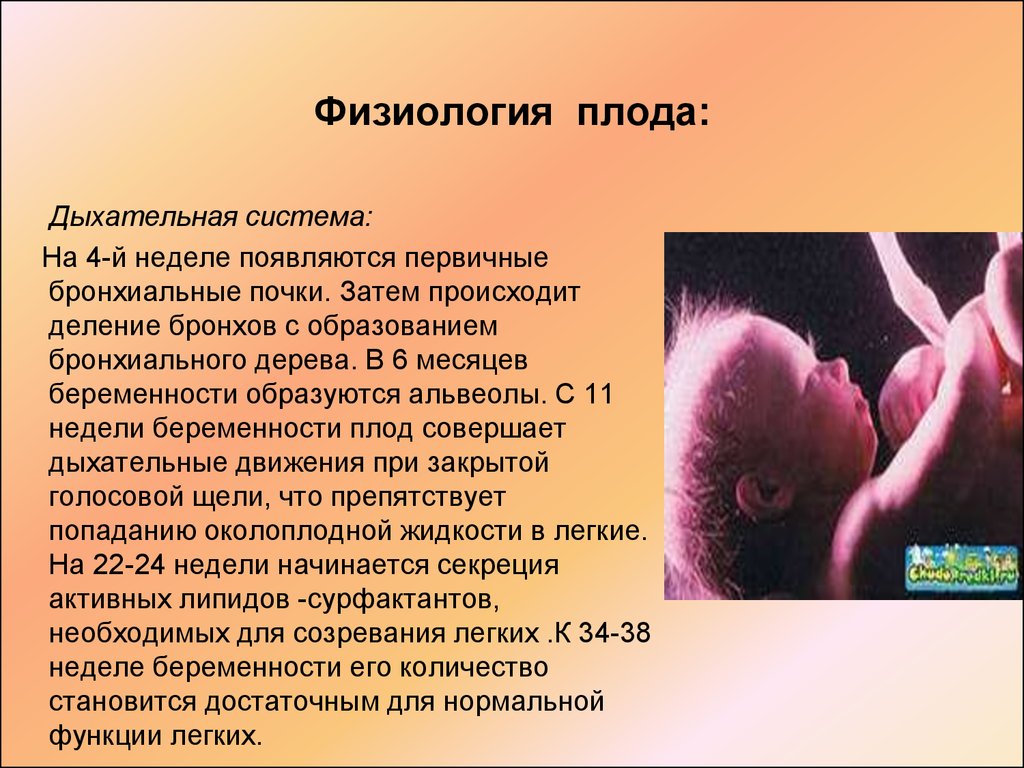
12-13 нед формир сурфактант, желточный мешок исчезает, четко определяются конечности, пальцырук и ног, появляются первые ядра окостенения, возможна визуализация четырехкамерного сердца.
К концу 16-й нед длина — 12 см, а масса тела — 100 г. 16 нед- оптимальное время ультразвукового скрининга для исключ патологии плода.
20
нед- длина плода — 25-26 см, масса 280-300 г.
Кожные покровы — красный цвет и обильно
покрыты пушковыми волосами и сыровидной
смазкой, являющейся продуктом деятельности
сальных желез. В кишечнике -образовывается
первородный кал (меконий).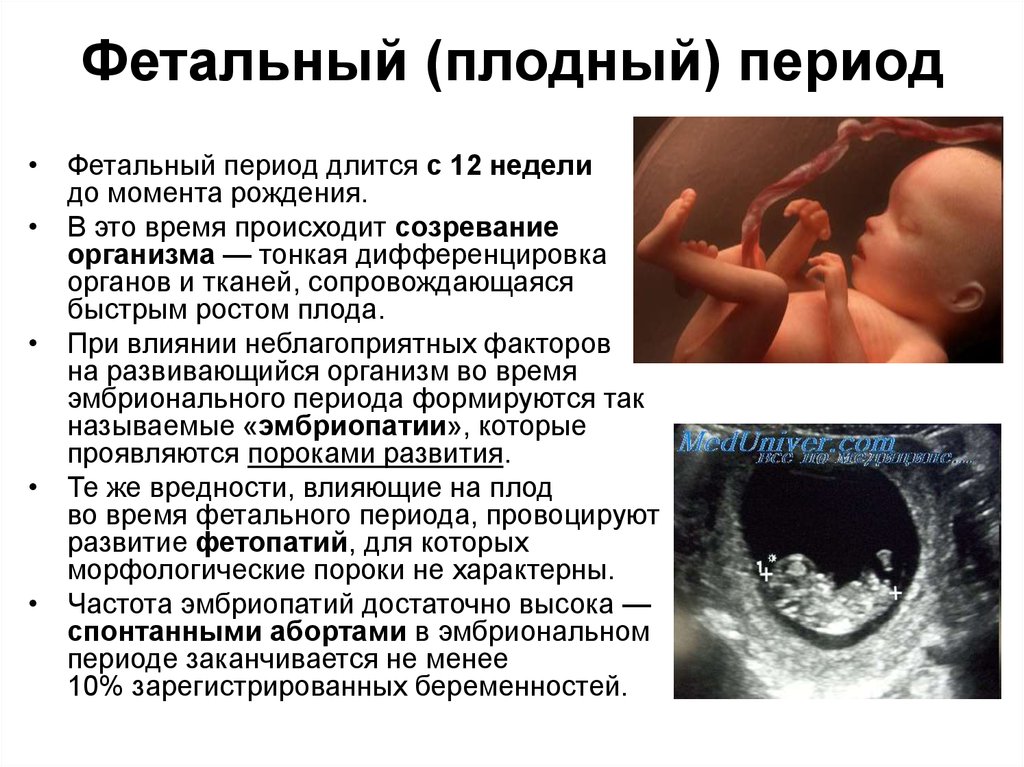 Первородящие
впервые начинают ощущать движения плода
(повторнородящие на 2 нед раньше). С
помощью акушерского стетоскопа — удается
впервые выслушать слабые сердцебиения
плода.
Первородящие
впервые начинают ощущать движения плода
(повторнородящие на 2 нед раньше). С
помощью акушерского стетоскопа — удается
впервые выслушать слабые сердцебиения
плода.
В конце VI(24нед) — длина плода — 30 см, масса 600-680 г. Такой плод может родиться живым и даже выжить при условии содержания его в специальных кювезах, при наличии ИВЛ и средств интенсивной терапии и реанимации.
В конце 7 месяца (28 нед) длина плода — 35 см, масса тела 1000-1200 г. У такого плода при рождении отмечаются выраженные признаки недоношенности: слабое развитие подкожной жировоЙ клетчатки, морщинистая кожа, обильная сыровидная смазка, выраженное развитие пушковых волос на теле. Хрящи носа и ушей мягкие, ногти не доходят до конца пальцев рук и ног. У мальчиков яички еще не опущены в мошонку, у девочек малые половые губы не прикрыты большими.
В конце 8 мес(32нед) 40-42 см, 1500-1700 г.
К концу 9 мес 45-48 см и 2400-2500 г.
В
конце 10 месяца у плода исчезают все
признаки недоношенности и имеются все
показатели, при сущие зрелому плоду.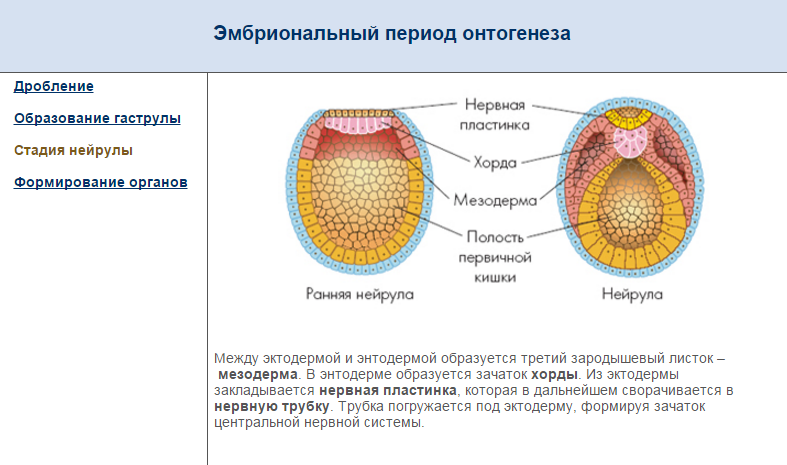
С 22-й недели внутриутробного развития, когда масса 500 г и более, — перинатальный период:последующие недели беременности+ роды+ 7 дней жизни новорожденного.
Критерии жизнеспособности по ВОЗ(минимальный шанс выжить): 22нед-500гр-25 см (шанс выжить-5%), 28нед-1кг,35см
Живорожденность-рождение с признаками жизни: крик, сердцебиение, спонтанные движения, пульсация пуповины
Зрелость новорожденного:
• грудь выпуклая, пупочное кольцо находится на середине между лобком и мечевидным отростком
• кожа бледно-роз цвета, ПЖК развита достаточно, пушковые волосы почти отсутствуют, длина волос на головке достигает 2см, ногти на ногах и руках доходят до кончиков пальцев;
• хрящи ушных раковин и носа упругие;
Физиология эмбрионального развития | SpringerLink
Аруши, Хурана И. Эмбриология человека. 1-е изд. Нью-Дели: CBS Publisher & Distributors Pvt Ltd.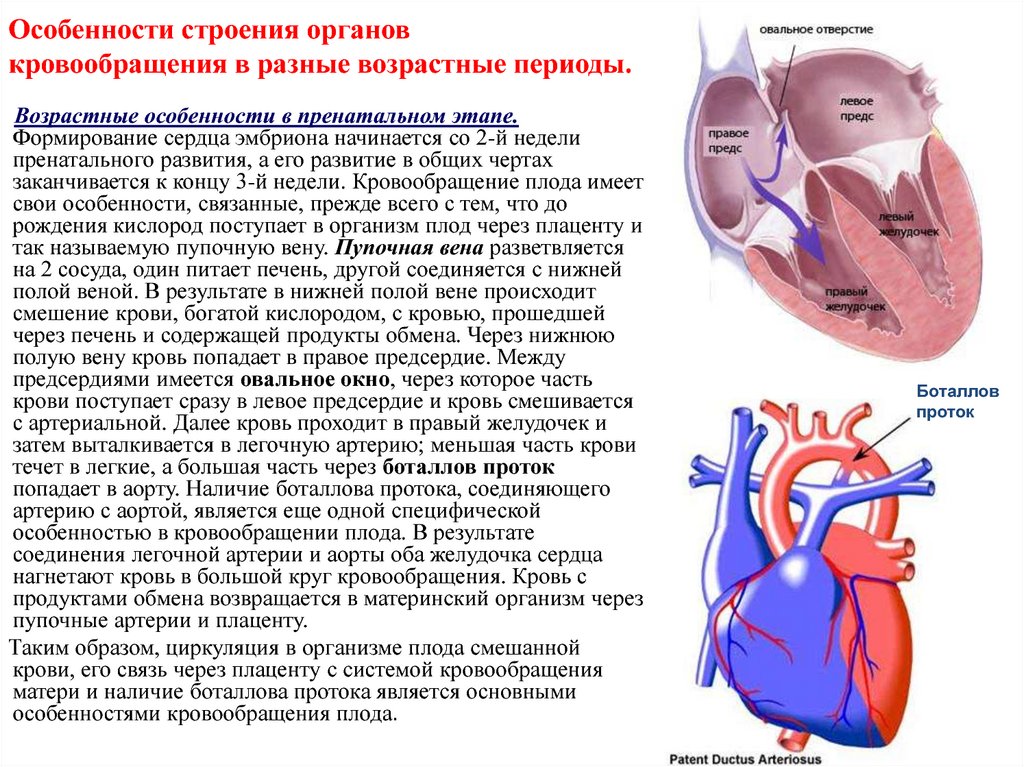 ; 2010.
; 2010.
Google ученый
Гуляс Б.Ж. Повторное исследование моделей дробления яиц плацентарных млекопитающих: вращение пар бластомеров во время второго дробления у кролика. Джей Эксп Зоол. 1975; 193: 235–48.
Перекрёстная ссылка КАС пабмед Google ученый
Гарднер Р.Л. Ранняя бластоциста билатерально симметрична, и ее ось симметрии совпадает с анимально-вегетативной осью зиготы мыши. Разработка. 1997; 124: 289–301.
КАС пабмед Google ученый
Гарнер В., Макларен А. Распределение клеток в химерных эмбрионах мышей перед имплантацией. J Embryol Exp Morphol. 1974; 32: 495–503.
КАС пабмед Google ученый
Беддингтон Р.С., Робертсон Э.Дж. Развитие оси и ранняя асимметрия у млекопитающих.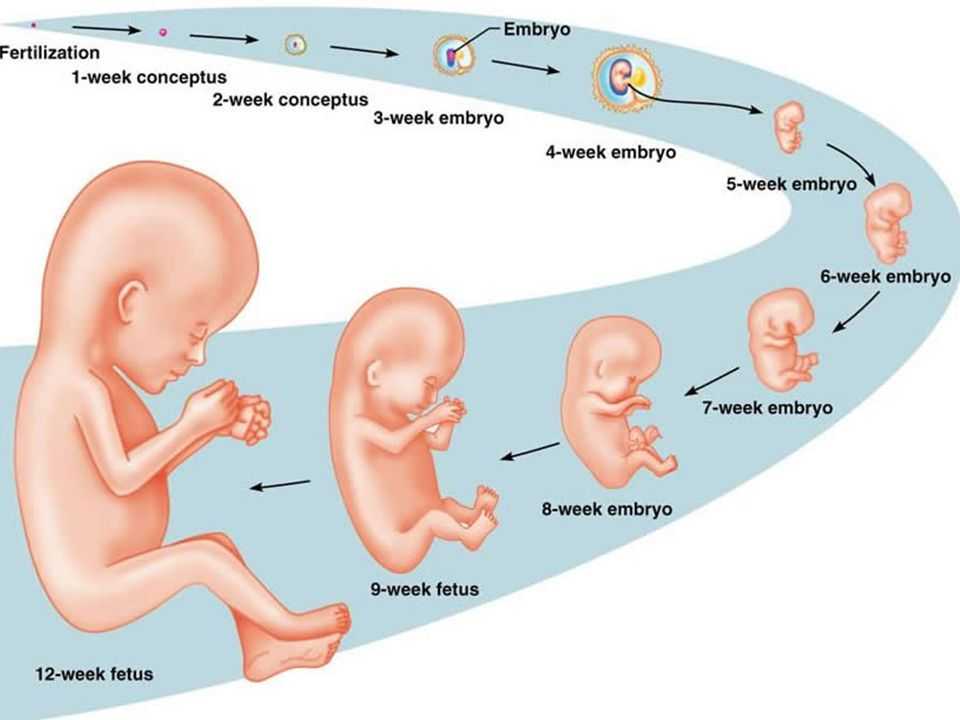 Клетка. 1999; 96: 195–209.
Клетка. 1999; 96: 195–209.
Перекрёстная ссылка КАС пабмед Google ученый
Гросс PR, Кузино Г.Х. Синтез белков, связанных с веретеном веретена, в раннем расщеплении. Джей Селл Биол. 1963; 19: 260–5.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google ученый
Кросби И.М., Гандольфи Ф., Мур Р.М. Контроль синтеза белка при раннем дроблении эмбрионов овцы. J Reprod Fertil. 1988; 82: 769–75.
Перекрёстная ссылка КАС пабмед Google ученый
Ли С., Гилула Н.Б., Уорнер А.Е. Щелевое соединение и уплотнение на предимплантационных стадиях развития мышей. Клетка. 1987; 51: 851–60.
Перекрёстная ссылка КАС пабмед Google ученый
 «>
«>Леви Дж. Б., Джонсон М. Х., Гудолл Х. и др. Время уплотнения: контроль основного перехода в развитии в раннем эмбриогенезе мышей. J Embryol Exp Morphol. 1986; 95: 213–37.
КАС пабмед Google ученый
Хэндисайд А.Х. Распределение сайтов связывания антител и лектинов на диссоциированных бластомерах мышиных морул: свидетельство поляризации при уплотнении. J Embryol Exp Morphol. 1980;60:99–116.
КАС пабмед Google ученый
Pratt HP, Ziomek CA, Reeve WJ, et al. Уплотнение эмбриона мыши: анализ его компонентов. J Embryol Exp Morphol. 1982; 70: 113–32.
КАС пабмед Google ученый
Рив В.Дж., Зиомек, Калифорния. Распределение микроворсинок на диссоциированных бластомерах эмбрионов мыши: свидетельство поляризации поверхности при уплотнении.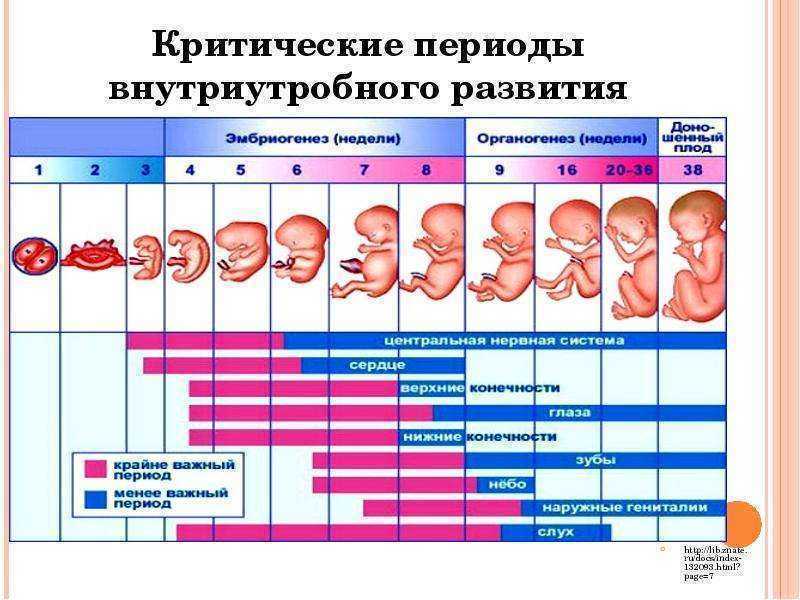 J Embryol Exp Morphol. 1981;62:339–350.
J Embryol Exp Morphol. 1981;62:339–350.
КАС пабмед Google ученый
Sutherland AE, Speed TP, Calarco PG. Распределение внутренних клеток в моруле мыши: роль ориентированного деления во время четвертого дробления. Дев биол. 1990; 137:13–25.
Перекрёстная ссылка КАС пабмед Google ученый
Карлсон Б.М. Основы эмбриологии. 6-е изд. Нью-Йорк: Макгроу-Хилл; 1996.
Google ученый
Барлоу П.В., Шерман Мичиган. Биохимия дифференцировки трофобласта мыши: исследования полиплоидии. J Embryol Exp Morphol. 1972; 27: 447–65.
КАС пабмед Google ученый
Джонсон М.Х., МакКоннелл Дж.М. Распределение клонов и клеточная полярность во время эмбриогенеза мыши.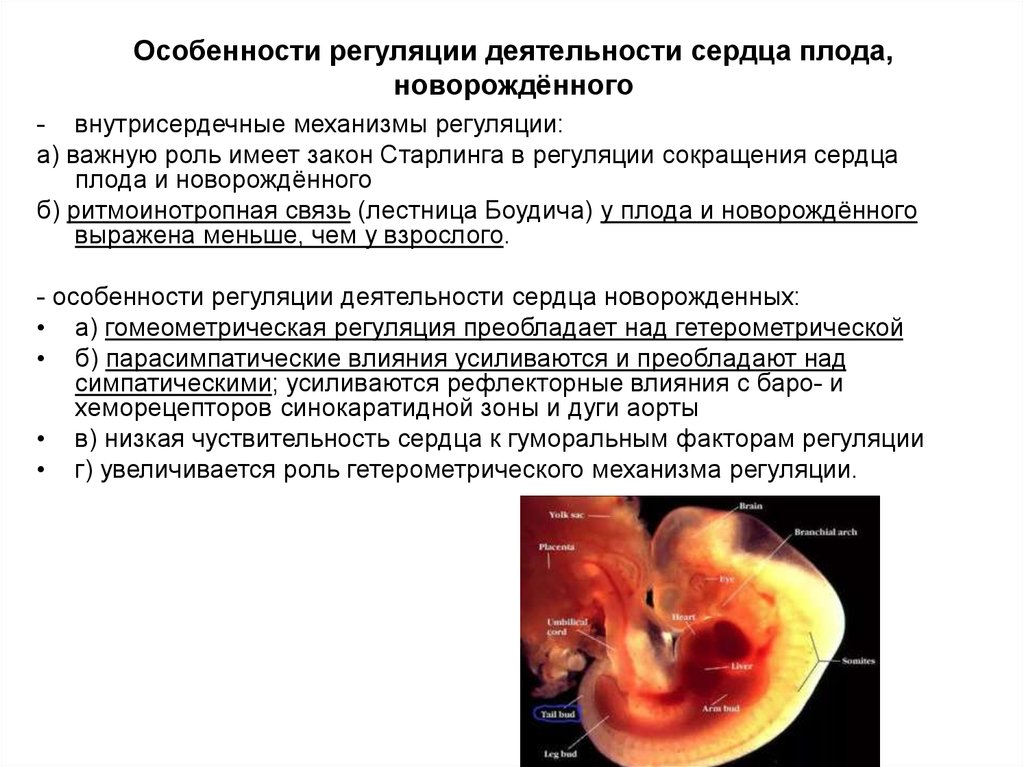 Semin Cell Dev Biol. 2004; 15: 583–97.
Semin Cell Dev Biol. 2004; 15: 583–97.
Перекрёстная ссылка КАС пабмед Google ученый
Марикава Ю., Аларкон В.Б. Установление линий трофэктодермы и внутренней клеточной массы у эмбриона мыши. Мол Репрод Дев. 2009;76:1019–32.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google ученый
Мюнтенер М., Хсу Ю.К. Развитие трофобласта и плаценты мыши. Повторное исследование культуры трофобласта и плаценты мыши in vitro. Акта Анат (Базель). 1977;98: 241–52.
Перекрёстная ссылка Google ученый
Флеминг ТП. Количественный анализ распределения клеток в трофэктодерме и внутренней клеточной массе бластоцисты мыши. Дев биол. 1987; 119: 520–31.
Перекрёстная ссылка КАС пабмед Google ученый
Pijnenborg R, Robertson WB, Brosens I, et al. Обзорная статья: инвазия трофобласта и формирование гемохориальной плаценты у человека и лабораторных животных. Плацента. 1981;2:71–91.
Перекрёстная ссылка КАС пабмед Google ученый
Ziomek CA, Johnson MH. Роль фенотипа и положения в управлении судьбой 16-клеточных бластомеров мыши. Дев биол. 1982; 91: 440–7.
Перекрёстная ссылка КАС пабмед Google ученый
Yamanaka Y, Ralston A, Stephenson RO, et al. Клеточная и молекулярная регуляция бластоцисты мыши. Дев Дин. 2006; 235:2301–14.
Перекрёстная ссылка КАС пабмед Google ученый
Гото Т., Монк М. Регуляция инактивации Х-хромосомы в развитии у мышей и людей. Microbiol Mol Biol Rev. 1998;62:362–78.
КАС ПабМед Центральный пабмед Google ученый
 «>
«>McGrath J, Solter D. Для завершения эмбриогенеза мыши необходимы как материнский, так и отцовский геномы. Клетка. 1984;37:179–83.
Перекрёстная ссылка КАС пабмед Google ученый
Гарбатт С.Л., Джонсон М.Х., Джордж Массачусетс. Когда и как порядок клеточного деления влияет на распределение клеток во внутренней клеточной массе бластоцисты мыши? Разработка. 1987; 100: 325–32.
КАС пабмед Google ученый
Гердон Дж. Б., Бирн Дж. А. Первые полвека ядерной трансплантации. Proc Natl Acad Sci U S A. 2003;100:8048–52.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google ученый
Смит Дж.М. Теория эволюции. Кембридж: Издательство Кембриджского университета; 1993.
Google ученый
 «>
«>Гиберт С.Ф. Биология развития. Сандерленд: Sinauer Associates, Inc.; 2000.
Google ученый
Уильямс ГК. Адаптация и естественный отбор. Принстон: Издательство Принстонского университета; 1996.
Google ученый
Гердон Дж. Б., Хопвуд Н. Введение Xenopus laevis в биологию развития: империя, тестирование на беременность и рибосомные гены. Int J Dev Biol. 2000; 44:43–50.
КАС пабмед Google ученый
Сандер К., Faessler PE. Представляем органайзер Шпемана-Мангольда: эксперименты и идеи, которые породили ключевую концепцию в биологии развития идеи, которые породили ключевую концепцию в биологии развития. Int J Dev Biol. 2001; 45:1–11.
КАС пабмед Google ученый
 «>
«>Вулперт Л., Джесселл Т., Лоуренс П. и др. Принципы развития. 3-е изд. Оксфорд: Издательство Оксфордского университета; 2007.
Google ученый
Дэвидсон Э.Х. Активность генов в раннем развитии. 2-е изд. Нью-Йорк: академический; 1976. с. 452.
Google ученый
Гандольфи Т.А., Гандольфи Ф. Материнское наследие эмбриону: цитоплазматические компоненты и их влияние на раннее развитие. Териогенология. 2001; 55: 1255–76.
Перекрёстная ссылка КАС пабмед Google ученый
Мемили Э., Первый NL. Зиготическая и эмбриональная экспрессия генов у коров: обзор сроков и механизмов ранней экспрессии генов по сравнению с другими видами. Зигота. 2000; 8: 87–96.
Перекрёстная ссылка КАС пабмед Google ученый
 «>
«>Биггерс Д.Д., Borland RM. Физиологические аспекты роста и развития преимплантационного эмбриона млекопитающих. Annu Rev Physiol. 1976; 38: 95–119.
Перекрёстная ссылка КАС пабмед Google ученый
Дворкин М.Б., Дворкин-Растл Э. Функции материнской мРНК в раннем развитии. Мол Репрод Дев. 1990; 26: 261–97.
Перекрёстная ссылка КАС пабмед Google ученый
Ван К., Чанг Ю.Г., де Врис В.Н., Струве М., Латам К.Е. Роль синтеза белка в развитии транскрипционно пермиссивного состояния у эмбрионов мышей на стадии одной клетки. Биол Репрод. 2001; 65: 748–54.
Перекрёстная ссылка КАС пабмед Google ученый
Бао С., Обата И., Кэрролл Дж. и др. Эпигенетические модификации, необходимые для нормального развития, устанавливаются при росте ооцитов у мышей. Биол Репрод. 2000;62:616–21.
Биол Репрод. 2000;62:616–21.
Перекрёстная ссылка КАС пабмед Google ученый
Аллегруччи С., Терстон А., Лукас Э. и др. Эпигенетика и зародышевая линия. Репродукция. 2005; 129:137–49.
Перекрёстная ссылка КАС пабмед Google ученый
Тада М., Тада Т., Лефевр Л. и др. Эмбриональные зародышевые клетки индуцируют эпигенетическое перепрограммирование соматического ядра в гибридных клетках. EMBO J. 1997; 16: 6510–20.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google ученый
Simon I, Tenzen T, Reubinoff BE, et al. Асинхронная репликация импринтированных генов устанавливается в гаметах и поддерживается во время развития. Природа. 1999; 401:929–32.
Перекрёстная ссылка КАС пабмед Google ученый
 «>
«>Хайкова П., Эрхардт С., Лейн Н. и др. Эпигенетическое перепрограммирование первичных зародышевых клеток мыши. Мех Дев. 2002; 117:15–23.
Перекрёстная ссылка КАС пабмед Google ученый
Obata Y, Kono T. Первичный импринтинг матери устанавливается в определенное время для каждого гена на протяжении всего роста ооцита. Дж. Биол. Хим. 2002; 277:5285–9.
Перекрёстная ссылка КАС пабмед Google ученый
Окано М., Белл Д.В., Хабер Д.А. и др. ДНК-метилтрансферазы Dnmt3a и Dnmt3b необходимы для метилирования de novo и развития млекопитающих. Клетка. 1999;99:247–57.
Перекрёстная ссылка КАС пабмед Google ученый
DiZio SM, Tasca RJ. Натрий-зависимый транспорт аминокислот у преимплантационных эмбрионов мышей: III. Na + -K + -ATPase-связанный механизм в бластоцистах. Дев биол. 1977; 59: 198–205.
Na + -K + -ATPase-связанный механизм в бластоцистах. Дев биол. 1977; 59: 198–205.
Перекрёстная ссылка КАС пабмед Google ученый
Йоханссон М., Янссон Т., Пауэлл Т.Л. Na(+)-K(+)-АТФаза распределяется в микроворсинках и базальной мембране синцитиотрофобласта плаценты человека. Am J Physiol Regul Integr Comp Physiol. 2000;279: R287–94.
КАС пабмед Google ученый
Ву Т.К., Лю Р.В., Хааксма С.Дж., Томасек Д.Дж. и др. Идентификация и клонирование ассоциированной с мембраной сериновой протеазы хепсина из преимплантационных эмбрионов мышей. Дж. Биол. Хим. 1997; 272:31315–20.
Перекрёстная ссылка КАС пабмед Google ученый
Перона Р.М., Вассарман П.М. Бластоцисты мыши вылупляются in vitro с помощью трипсиноподобной протеиназы, связанной с клетками пристеночной трофэктодермы.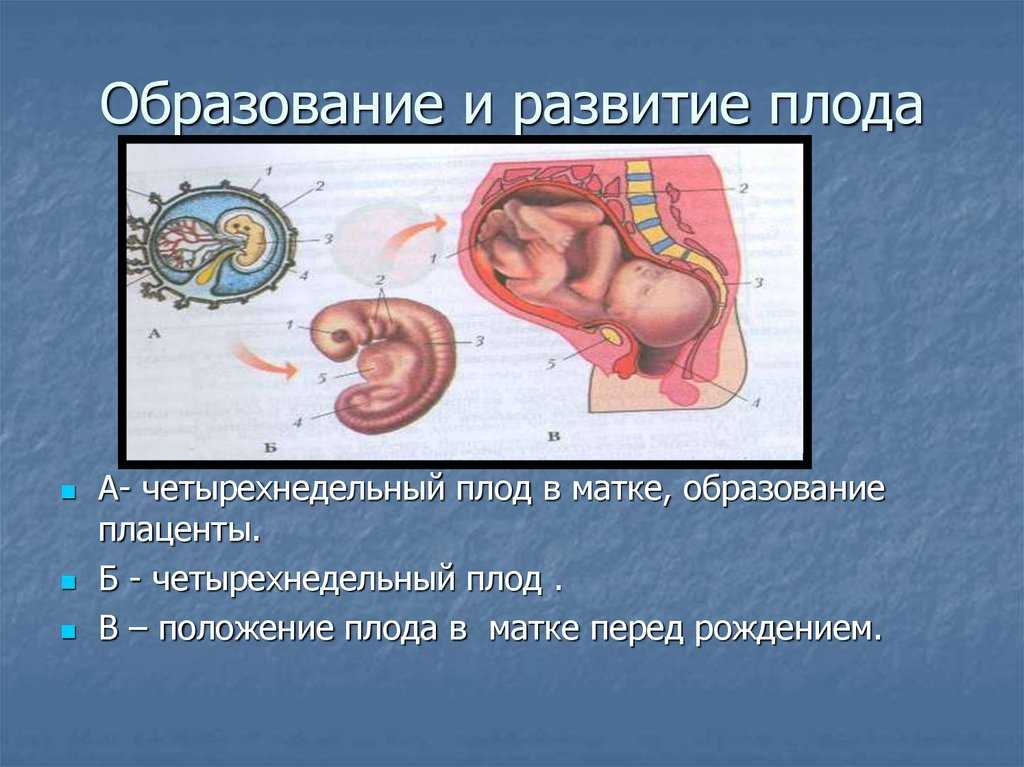 Дев биол. 1986;114:42–52.
Дев биол. 1986;114:42–52.
Перекрёстная ссылка КАС пабмед Google ученый
Das SK, Wang XN, Paria BC и др. Гепарин-связывающий ген EGF-подобного фактора роста индуцируется в матке мышей бластоцистой височно только в месте ее прилегания: возможный лиганд для взаимодействия с EGF-рецептором бластоцисты при имплантации. Разработка. 1994; 120:1071–83.
КАС пабмед Google ученый
Aplin JD, Seif MW, Graham RA, et al. Поверхность клеток эндометрия и имплантация. Экспрессия полиморфного муцина MUC-1 и молекул адгезии во время эндометриального цикла. Энн Н.Ю. Академия наук. 1994; 734: 103–21.
Перекрёстная ссылка КАС пабмед Google ученый
Сидху С.С., Кимбер С.Дж. Гормональный контроль альфа-(1–2)фукозилтрансферазы Н-типа мессенджера рибонуклеиновой кислоты в матке мышей. Биол Репрод. 1999;60(1):147–57.
Биол Репрод. 1999;60(1):147–57.
Перекрёстная ссылка КАС пабмед Google ученый
Гилберт С.Ф. Эпидермис и происхождение кожных структур. В: Биология развития. 6-е изд. Сандерленд: Sinauer Associates; 2000.
Google ученый
Гилберт С.Ф. Сравнительная эмбриология. В: Биология развития. 6-е изд. Сандерленд: Sinauer Associates; 2000.
Google ученый
Гилберт С.Ф. Раннее развитие млекопитающих. В: Биология развития. 6-е изд. Сандерленд: Sinauer Associates; 2000.
Google ученый
Холл БК. Нервный гребень как четвертый зародышевый слой и позвоночные как квадробластный, а не триплобластный. Эвол Дев. 2000; 2: 3–5.
Перекрёстная ссылка КАС пабмед Google ученый
 «>
«>Мур КЛ. Развивающийся человек. 2-е изд. Филадельфия: Сондерс; 1977.
Google ученый
Мур КЛ. Развивающийся человек: клинически ориентированная эмбриология. 4-е изд. Филадельфия: Сондерс; 1988.
Google ученый
Мур КЛ. Прежде чем мы родимся. Базовая эмбриология и врожденные дефекты. Филадельфия: Сондерс; 1983.
Google ученый
Usher R, Shephard M, Lind J. Объем крови новорожденного и переливание плаценты. Акта Педиатр. 1963;52:497–512.
Перекрёстная ссылка КАС пабмед Google ученый
Янссон Т., Пауэлл Т.Л. Плацентарный транспорт человека при измененном росте плода: функционирует ли плацента как датчик питательных веществ? Обзор. Плацента. 2006;27:S91.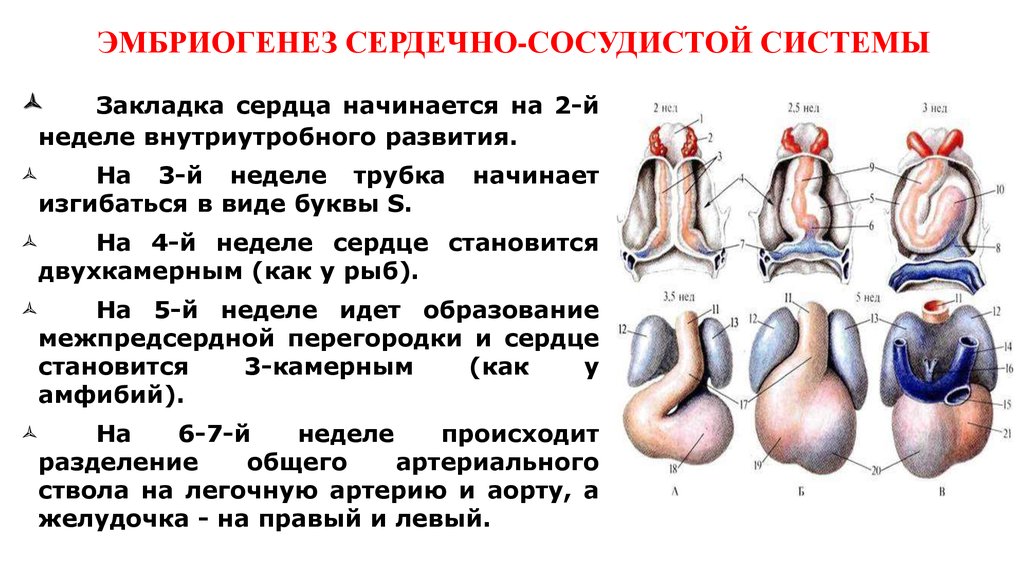
Перекрёстная ссылка пабмед Google ученый
Sipes SL, Weiner CP, Wenstrom KD, et al. Связь между кариотипом плода и средним корпускулярным объемом. Am J Obstet Gynecol. 1991;165:1371–1376.
Перекрёстная ссылка КАС пабмед Google ученый
Пирсон HA. Последние достижения гематологии. J Педиатр. 1966; 69: 466–79.
Перекрёстная ссылка КАС пабмед Google ученый
Weiner CP, Sipes SL, Wenstrom K. Влияние возраста плода на нормальные лабораторные показатели плода и венозное давление. Акушерство Гинекол. 1992; 79: 713–18.
КАС пабмед Google ученый
Фрайер А.А., Джонс П., Стрэндж Р. и др. Уровни белка плазмы у нормальных плодов человека: от 13 до 41 недели беременности.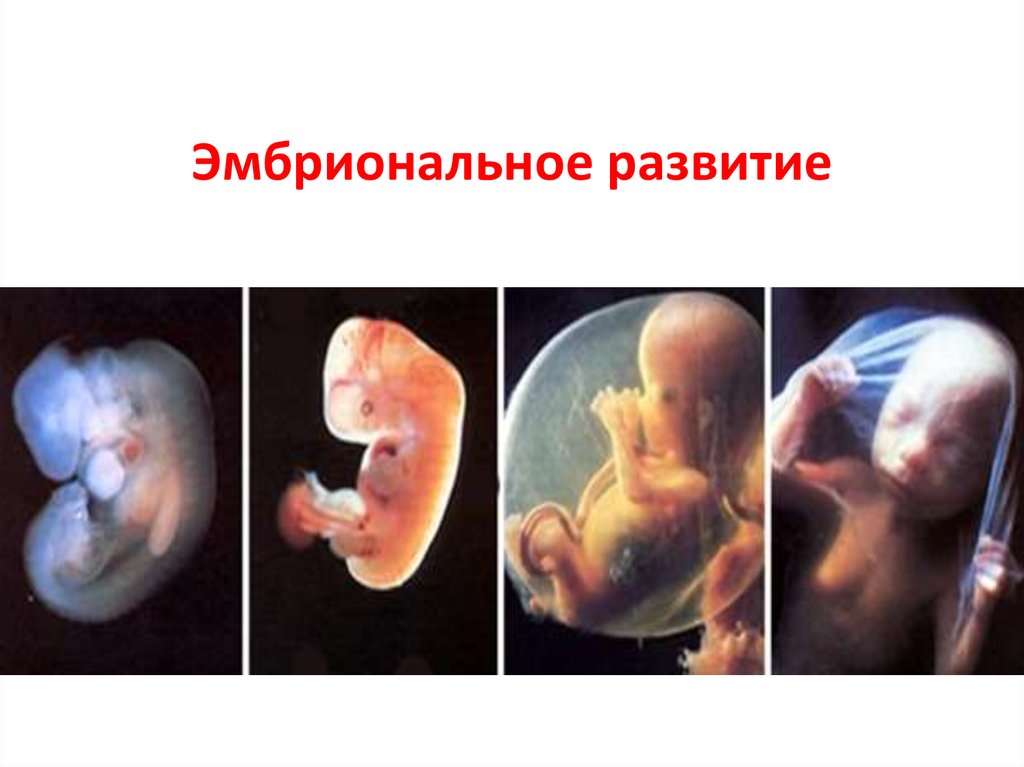 Br J Obstet Gynaecol. 1993; 100:850–5.
Br J Obstet Gynaecol. 1993; 100:850–5.
Перекрёстная ссылка КАС пабмед Google ученый
Foley ME, Isherwood DM, McNicol GP. Вязкость, гематокрит, фибриноген и белки плазмы в материнской и пуповинной крови. Br J Obstet Gynaecol. 1978; 85: 500–4.
Перекрёстная ссылка КАС пабмед Google ученый
Колдовский О., Херингова А., Жирсова В. и др. Транспорт глюкозы против градиента концентрации в вывернутых мешочках тощей и подвздошной кишки плодов человека. Гастроэнтерология. 1965; 48: 185–7.
КАС пабмед Google ученый
Миллер А.Дж. Глотание. Physiol Rev. 1982; 62: 129–84.
КАС пабмед Google ученый
Притчард Дж.А. Глотание плода и объем амниотической жидкости. Акушерство Гинекол. 1966;28:606–10.
Акушерство Гинекол. 1966;28:606–10.
КАС пабмед Google ученый
Лебенталь Э., Ли П.С. Обзорная статья. Взаимодействия детерминант онтогенеза желудочно-кишечного тракта: единая концепция. Педиатр Рез. 1983; 1: 19–24.
Перекрёстная ссылка Google ученый
Башор Р.А., Смит Ф., Шенкер С. Плацентарный перенос и распределение билирубина у беременной обезьяны. Am J Obstet Gynecol. 1969;103:950–8.
КАС пабмед Google ученый
Адам П.Дж., Терамо К., Райха Н. и др. Метаболизм инсулина плода человека на ранних сроках беременности: реакция на острое повышение концентрации глюкозы плода и плацентарный перенос человеческого инсулина-I-131. Диабет. 1969; 18: 409–16.
Перекрёстная ссылка КАС пабмед Google ученый
 «>
«>Обеншайн С.С., Адам П.Дж., Кинг К.С. и др. Реакция инсулина плода человека на устойчивую материнскую гипергликемию. N Engl J Med. 1970;283:566–70.
Перекрёстная ссылка КАС пабмед Google ученый
Верлин С.Л. Экзокринная поджелудочная железа. В: Полин Р.А., Фокс В.В., ред. Физиология плода и новорожденного. Филадельфия: Сондерс; 1992. с. 1047.
Google ученый
Davis MM, Hodes ME, Munsick RA, et al. Экспрессия панкреатической амилазы в развитии поджелудочной железы человека. Гибридома. 1986; 5: 137–45.
перекрестная ссылка КАС пабмед Google ученый
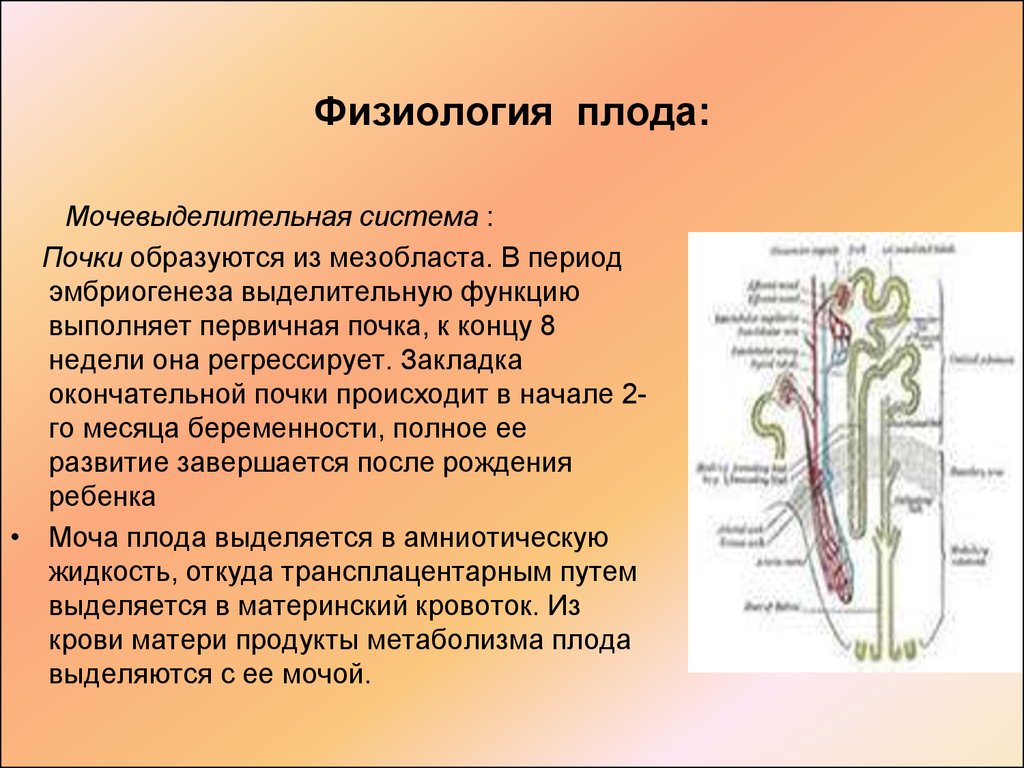
Саксен Л., Сариола Х. Ранний органогенез почки. Педиатр Нефрол. 1987; 1: 385–92.
Перекрёстная ссылка пабмед Google ученый
 «>
«>Geelhoed JJ, Verburg BO, Nauta J, et al. Отслеживание и детерминанты размера почек от внутриутробной жизни до 2-летнего возраста: исследование поколения R. Ам Джей Почка. 2009;53(2):248–58.
Перекрёстная ссылка Google ученый
Smith FG, Nakamura KT, Segar JL et al. В: Полин Р.А., Фокс В.В., ред. Физиология плода и новорожденного, т. 2, гл. 114. Филадельфия: Сондерс; 1992. с. 1187.
Google ученый
Wladimiroff JW, Campbell S. Нормы продукции мочи плода при нормальной и осложненной беременности. Ланцет. 1974; 1: 151–4.
Перекрёстная ссылка КАС пабмед Google ученый
Chard T, Hudson CN, Edwards CRW, et al. Высвобождение окситоцина и вазопрессина плодом человека во время родов. Природа. 1971; 234: 352–4.
Перекрёстная ссылка КАС пабмед Google ученый
 «>
«>Полин Р.А., Хусейн М.К., Джеймс Л.С. и др. Высокие концентрации вазопрессина в пуповинной крови человека — отсутствие корреляции со стрессом. J Перинат Мед. 1977; 5: 114–19.
Перекрёстная ссылка КАС пабмед Google ученый
Баллабио М., Николини У., Джоуэтт Т. и др. Созревание функции щитовидной железы у нормальных плодов человека. Клин Эндокринол. 1989; 31: 565–71.
Перекрёстная ссылка КАС Google ученый
Thorpe-Beeston JG, Nicolaides KH, Felton CV, et al. Созревание секреции гормонов щитовидной железы и тиреотропного гормона у плода. N Engl J Med. 1991; 324: 532–6.
Перекрёстная ссылка КАС пабмед Google ученый
Wenstrom KD, Weiner CP, Williamson RA, et al. Пренатальная диагностика гипертиреоза плода с помощью фунипунктуры. Акушерство Гинекол. 1990; 76: 513–17.
Акушерство Гинекол. 1990; 76: 513–17.
КАС пабмед Google ученый
Вулсма Т., Гонс М.Х., Де Вийлдер Дж.Дж. Трансфер тироксина от матери к плоду при врожденном гипотиреозе вследствие тотального дефекта организации или агенезии щитовидной железы. N Engl J Med. 1989; 321:13–6.
Перекрёстная ссылка КАС пабмед Google ученый
Кофф А.К. Развитие влагалища у плода человека. Внести Эмбриол. 1933; 24: 59–91.
КАС пабмед Google ученый
Кониси И., Фуджи С., Окамура Х. и др. Развитие интерстициальных клеток и яйцеклеток в яичниках плода человека: ультраструктурное исследование. Дж Анат. 1986; 148: 121–35.
КАС ПабМед Центральный пабмед Google ученый
 «>
«>Bozzetti P, Ferrari MM, Marconi AM и др. Взаимосвязь концентраций глюкозы матери и плода у человека с середины беременности до срока. Метаболизм. 1988; 37: 358–63.
Перекрёстная ссылка КАС пабмед Google ученый
Hauguel-de Mouzon S, Lepercq J, Catalano P. Известное и неизвестное о лептине во время беременности. Am J Obstet Gynecol. 2006;193(6):1537–45.
Перекрёстная ссылка Google ученый
Grisaru-Granovsy S, Samueloff A, Elstein D. Роль лептина в росте плода: краткий обзор от зачатия до родов. Eur J Obstet Gynecol Reprod Biol. 2008;136(2):146–50.
Перекрёстная ссылка Google ученый
Кимура Р.Э. Липидный обмен в плодово-плацентарном комплексе. В: Коуэтт Р.М., редактор. Принципы перинатально-неонатального метаболизма. Нью-Йорк: Спрингер; 1991. с. 291.
Нью-Йорк: Спрингер; 1991. с. 291.
Google ученый
Лимоны JA. Метаболизм азота в плаценте плода. Семин Перинатол. 1979; 3: 177–90.
КАС пабмед Google ученый
Morriss FH Jr, Boyd RDH, Manhendren D. Транспорт плаценты. В: Knobil E, Neill J, редакторы. Физиология размножения, том II. Нью-Йорк: Рэйвен; 1994. с. 813.
Google ученый
Fowden AL, Ward JW, Wooding FP, et al. Программирование способности плаценты транспортировать питательные вещества. Дж. Физиол. 2006;572(1):5–15.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google ученый
Янссон Т., Пауэлл Т.Л. Премия IFPA 2005 за лекцию по плацентологии. Плацентарный транспорт человека при измененном росте плода: функционирует ли плацента как датчик питательных веществ? — Обзор. Плацента. 2006; 27:S91–7.
Плацента. 2006; 27:S91–7.
Перекрёстная ссылка пабмед Google ученый
Гитлин Д., Кумате Дж., Моралес С. и др. Круговорот белков амниотической жидкости в зачатке человека. Am J Obstet Gynecol. 1972;113:632–45.
КАС пабмед Google ученый
Abbas SK, Pickard DW, Illingworth D, et al. Измерение белка ПТГ-рФ в экстрактах паращитовидных желез плода и плацентарных мембран. J Эндокринол. 1990; 124:319–25.
Перекрёстная ссылка КАС пабмед Google ученый
Хеллман П., Райдефельт П., Юхлин С. и др. Паратироидоподобная регуляция высвобождения белка, связанного с паратиреоидным гормоном, и цитоплазматического кальция в клетках цитотрофобласта плаценты человека. Арх Биохим Биофиз. 1992;293:174–80.
Перекрёстная ссылка КАС пабмед Google ученый
 «>
«>Гилберт В.М., Брейс Р.А. Объем амниотической жидкости и нормальные потоки в амниотическую полость и из нее. Семин Перинатол. 1993; 17: 150–7.
КАС пабмед Google ученый
Брейс Р.А., Вольф Э.Дж. Нормальный объем амниотической жидкости меняется на протяжении всей беременности. Я акушер-гинекол. 1989; 161: 382–8.
перекрестная ссылка КАС Google ученый
эмбрионов | человек и животное
первые этапы развития человека
Смотреть все СМИ
- Ключевые люди:
- Хван Ву-Сук Мун Шин Юн Ёсинори Осуми Этьен Жоффруа Сент-Илер сэр Джулиан Хаксли
- Связанные темы:
- половая дифференциация гестационный возраст имплантация расщепление сегментация
См. все связанные материалы →
эмбрион , ранняя стадия развития животного, пока оно находится в яйцеклетке или в матке матери. У людей этот термин применяется к нерожденному ребенку до конца седьмой недели после зачатия; с восьмой недели нерожденный ребенок называется плодом.
У людей этот термин применяется к нерожденному ребенку до конца седьмой недели после зачатия; с восьмой недели нерожденный ребенок называется плодом.
Далее следует краткое описание эмбрионального развития. Для полной обработки см. Морфология: Эмбриология.
Подробнее по этой теме
Морфология: Эмбриология
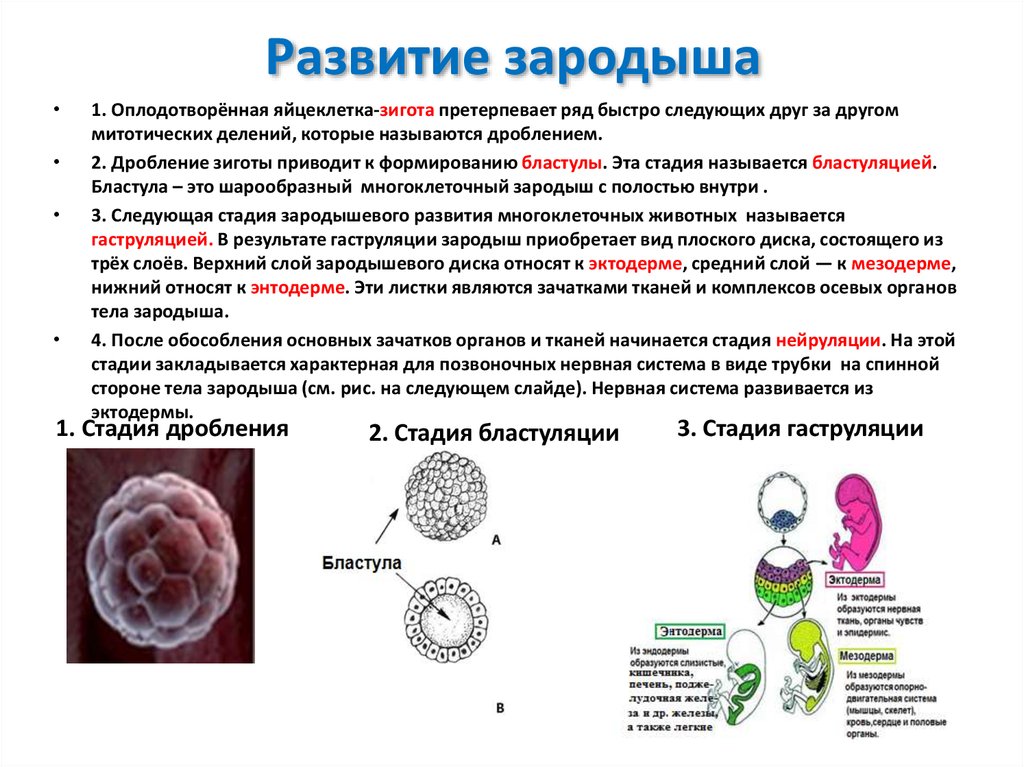
Развитие обычно начинается у животных с дробления или деления оплодотворенной яйцеклетки (зиготы) с образованием полого шара клеток, называемого…
У организмов, размножающихся половым путем, соединение яйцеклетки со сперматозоидом приводит к образованию зиготы или оплодотворенной яйцеклетки, которая проходит серию делений, называемых дроблением, по мере прохождения по фаллопиевой трубе. После нескольких дроблений клетки образуют полый шар, называемый бластулой. У большинства млекопитающих бластула прикрепляется к слизистой оболочке матки, стимулируя тем самым образование плаценты, которая будет передавать питательные вещества от матери к растущему эмбриону. У низших животных зародыш питается желтком.
У низших животных зародыш питается желтком.
В процессе гаструляции эмбрион дифференцируется на три типа тканей: эктодерму, производящую кожу и нервную систему; мезодерма, из которой развиваются соединительные ткани, кровеносная система, мышцы и кости; и энтодерма, которая формирует пищеварительную систему, легкие и мочевыделительную систему. Мезодермальные клетки мигрируют с поверхности эмбриона, чтобы заполнить пространство между двумя другими тканями через удлиненное углубление, известное как первичная полоска. По мере развития эмбриона слои клеток складываются, так что энтодерма образует длинную трубку, окруженную мезодермой, с эктодермальным слоем вокруг всего.
Питательные вещества проходят из плаценты через пуповину, а амнион, заполненная жидкостью оболочка, окружает и защищает эмбрион. Становится очевидным разделение тела на голову и туловище, начинают развиваться головной, спинной мозг и внутренние органы. Все эти изменения завершаются в начале эмбрионального развития, примерно к четвертой неделе у человека.