Содержание
Первые признаки: Как распознать 5 мм плодное яйцо на УЗИ
Беременность — удивительное время в жизни женщины, полное волнений и ожиданий. Но как узнать, что новая жизнь действительно начала свой путь? Современная медицина предлагает нам множество способов, и один из самых ранних и точных — это ультразвуковое исследование. Но что же мы можем увидеть на таком раннем сроке? Давайте разберемся, что такое 5 мм плодное яйцо и как его распознать.
Плодное яйцо — это, по сути, первое «жилище» будущего малыша. Оно формируется вскоре после того, как оплодотворенная яйцеклетка прикрепляется к стенке матки. Внутри этого крошечного пузырька и начинается волшебный процесс развития новой жизни. Но когда же его можно увидеть?
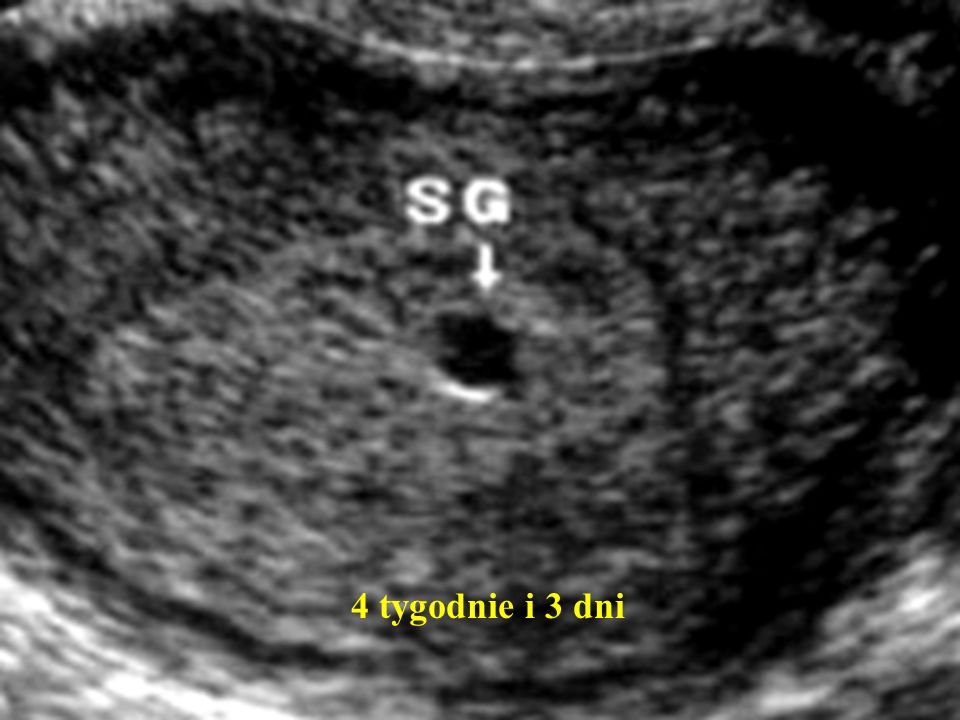
Обычно плодное яйцо размером 5 мм можно обнаружить на УЗИ примерно на 4-5 неделе беременности. Это невероятно ранний срок, не правда ли? Многие женщины в это время еще даже не подозревают о своем «интересном положении». Но для опытного врача-узиста такое плодное яйцо — уже весомое доказательство начавшейся беременности.
Как же выглядит 5 мм плодное яйцо на экране УЗИ? Представьте себе крошечный темный кружок или овал в море серого. Это и есть наше плодное яйцо! Оно выглядит как небольшая полость, заполненная жидкостью, внутри которой пока еще сложно что-либо разглядеть. Но не стоит расстраиваться — ведь это только начало удивительного путешествия!
Интересно, что размер плодного яйца в 5 мм — это своеобразная «отправная точка» для многих врачей. До этого момента беременность считается очень ранней и ее не всегда можно достоверно подтвердить на УЗИ. Но как только плодное яйцо достигает этого размера, шансы на успешное развитие беременности значительно возрастают.
Что можно увидеть внутри 5 мм плодного яйца?
На этом этапе внутри плодного яйца уже происходят важные процессы, хотя разглядеть их пока сложно. Эмбрион еще слишком мал — его размер составляет всего около 1-2 мм. Но не стоит недооценивать эту крошечную жизнь! Уже сейчас формируются важнейшие органы и системы будущего человека.

Внутри 5 мм плодного яйца можно увидеть так называемый желточный мешок — временный орган, который обеспечивает питание эмбриона на ранних стадиях развития. Он выглядит как маленький светлый кружок внутри плодного яйца. Наличие желточного мешка — хороший признак, говорящий о нормальном развитии беременности.
Иногда при очень тщательном исследовании можно заметить и крошечную точку — это и есть сам эмбрион. Но не стоит расстраиваться, если его пока не видно — на таком раннем сроке это абсолютно нормально. Главное — наличие самого плодного яйца и его правильная форма.
Как определить, что перед нами именно 5 мм плодное яйцо?
Определение размера плодного яйца — задача для профессионала. Врач-узист использует специальные инструменты для измерения, учитывая не только диаметр, но и объем плодного яйца. Важно помнить, что 5 мм — это средний показатель, и небольшие отклонения в ту или иную сторону вполне допустимы.
Кроме размера, врач обращает внимание на форму плодного яйца. Идеальная форма — правильный овал или круг. Если плодное яйцо имеет неправильную форму или его контуры нечеткие, это может быть поводом для беспокойства и требует дополнительного наблюдения.
Важно также оценить расположение плодного яйца в матке. Нормальное плодное яйцо должно располагаться в верхней части полости матки, примерно посередине. Если оно находится слишком низко или сбоку, это может указывать на проблемы с имплантацией.
Что делать, если на УЗИ обнаружено 5 мм плодное яйцо?
Обнаружение 5 мм плодного яйца на УЗИ — это отличная новость! Это значит, что беременность развивается, и малыш уже начал свой путь. Однако не стоит расслабляться — впереди еще долгий путь.
После обнаружения плодного яйца врач обычно назначает повторное УЗИ через 1-2 недели. За это время плодное яйцо должно значительно увеличиться в размерах, а внутри него уже можно будет разглядеть эмбрион и, возможно, даже услышать сердцебиение!
Важно помнить, что каждая беременность уникальна, и темпы развития могут немного отличаться. Не стоит паниковать, если на первом УЗИ плодное яйцо оказалось чуть меньше или больше 5 мм. Главное — его наличие и правильная форма.
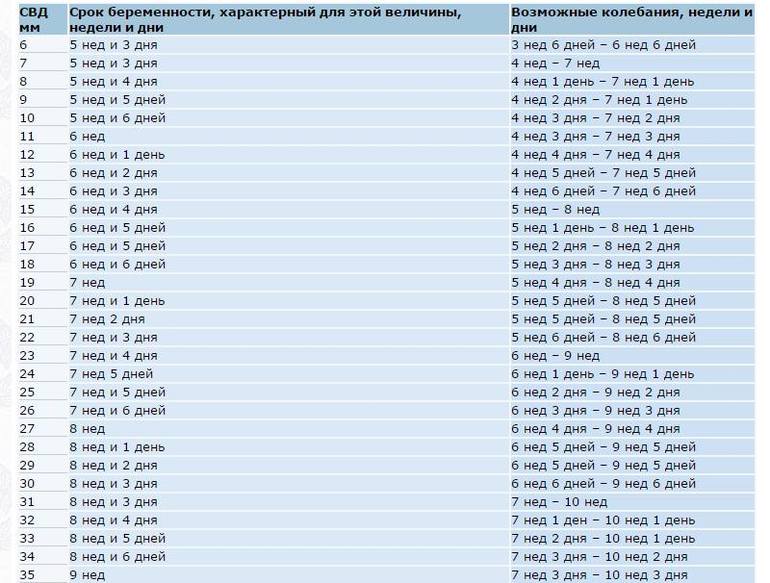
Можно ли определить срок беременности по размеру плодного яйца?
Размер плодного яйца действительно может помочь в определении срока беременности, особенно на ранних этапах. Однако важно понимать, что это не единственный и не самый точный метод.
Плодное яйцо размером 5 мм обычно соответствует сроку беременности около 4-5 недель от первого дня последней менструации. Но опять же, это средний показатель, и индивидуальные отклонения вполне возможны.
Для более точного определения срока беременности врачи используют комплексный подход, учитывая не только размер плодного яйца, но и другие параметры: дату последней менструации, уровень гормонов, общее состояние женщины.
Что может пойти не так?
Другая потенциальная проблема — это внематочная беременность. Иногда плодное яйцо может имплантироваться не в матке, а в маточной трубе. Это опасное состояние, требующее немедленного медицинского вмешательства.
Поэтому так важно регулярное наблюдение у врача и выполнение всех рекомендаций. Только так можно обеспечить здоровое развитие беременности и вовремя выявить возможные проблемы.
Заключительные мысли
Обнаружение 5 мм плодного яйца на УЗИ — это волнующий момент для будущих родителей. Это первое реальное доказательство того, что новая жизнь начала свой путь. Но помните, что это только начало удивительного путешествия под названием «беременность».
Впереди еще много важных этапов и открытий. Уже совсем скоро вы услышите сердцебиение своего малыша, увидите его первые движения на экране УЗИ, почувствуете его толчки. А пока — наслаждайтесь каждым моментом, заботьтесь о себе и не забывайте регулярно посещать врача. Ведь здоровье мамы — залог здоровья малыша!
Эмбриональное развитие: От оплодотворения до формирования плодного яйца
Мир эмбрионального развития — это настоящее чудо природы, где каждый день происходят удивительные метаморфозы. Представьте себе, что вы отправляетесь в микроскопическое путешествие, наблюдая за тем, как из двух крошечных клеток постепенно формируется новая жизнь. Это захватывающий процесс, который начинается с момента оплодотворения и продолжается вплоть до формирования плодного яйца размером 5 мм — первого видимого признака беременности на УЗИ.
Все начинается с встречи двух клеток — сперматозоида и яйцеклетки. Эта встреча происходит в маточной трубе, и с этого момента начинается удивительный танец жизни. Оплодотворенная яйцеклетка, теперь называемая зиготой, начинает свое путешествие к матке, где ей предстоит «поселиться» на ближайшие 9 месяцев.
По пути зигота активно делится, превращаясь сначала в морулу (напоминающую по форме ягоду ежевики), а затем в бластоцисту — полый шарик из клеток. Именно бластоциста и станет тем самым плодным яйцом, которое мы увидим на УЗИ через несколько недель.
Имплантация: Момент истины
Примерно на 6-7 день после оплодотворения бластоциста достигает матки и начинает процесс имплантации. Это критический момент — бластоциста должна «зацепиться» за стенку матки и начать формировать связи с материнским организмом. Процесс имплантации занимает несколько дней и сопровождается сложными биохимическими процессами.
Интересно, что именно в этот момент некоторые женщины могут заметить легкое кровотечение, известное как имплантационное. Оно часто принимается за начало менструации, но на самом деле является первым признаком наступившей беременности. Вот так природа играет с нами в прятки!
От бластоцисты к эмбриону
После успешной имплантации начинается активное развитие эмбриона. Клетки бластоцисты дифференцируются, образуя три зародышевых листка: эктодерму, мезодерму и энтодерму. Из этих трех слоев в будущем сформируются все органы и ткани малыша. Параллельно формируется плацента — уникальный орган, который будет обеспечивать связь между матерью и плодом на протяжении всей беременности.
К концу второй недели после оплодотворения (что соответствует примерно 4 неделям беременности по акушерскому сроку) формируется примитивная полоска — структура, определяющая ось тела будущего эмбриона. Это важнейший этап развития, после которого эмбрион начинает приобретать узнаваемые черты.
5 мм плодное яйцо: Первая веха
И вот мы подошли к ключевому моменту нашего путешествия — формированию плодного яйца размером 5 мм. Это происходит примерно на 4-5 неделе беременности (или 2-3 неделе после оплодотворения). Плодное яйцо в этот момент представляет собой небольшую полость, заполненную жидкостью, внутри которой находится крошечный эмбрион.
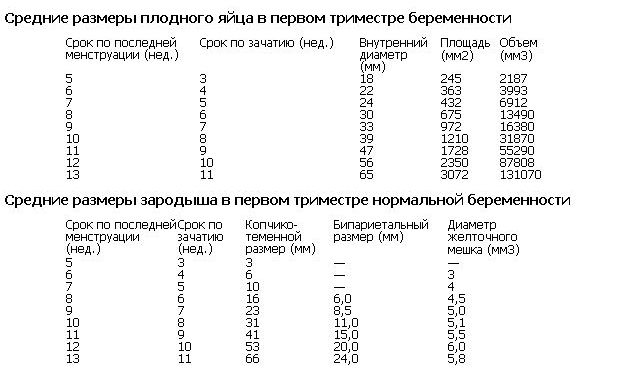
5 мм плодное яйцо — это своеобразный рубеж в развитии беременности. До этого момента беременность считается очень ранней, и ее не всегда можно достоверно подтвердить на УЗИ. Но как только плодное яйцо достигает размера 5 мм, оно становится видимым при ультразвуковом исследовании, что позволяет врачам с уверенностью диагностировать беременность.
Что происходит внутри 5 мм плодного яйца?
Несмотря на свои крошечные размеры, внутри 5 мм плодного яйца происходят грандиозные события. Эмбрион в это время имеет размер всего 1-2 мм, но уже начинает формировать основные органы и системы.
- Нервная трубка, из которой в будущем разовьются головной и спинной мозг, уже начала формироваться.
- Закладывается примитивное сердце, которое вскоре начнет биться.
- Формируются зачатки будущих конечностей.
- Начинает развиваться пищеварительная система.
Кроме того, внутри плодного яйца можно увидеть желточный мешок — временный орган, обеспечивающий питание эмбриона на ранних стадиях развития. Он выглядит как маленький светлый кружок внутри плодного яйца.
Как распознать 5 мм плодное яйцо на УЗИ?
На экране УЗИ 5 мм плодное яйцо выглядит как небольшой темный кружок или овал. Его размер измеряется от одного края до другого. Важно отметить, что на этом сроке сам эмбрион обычно еще не виден — он слишком мал даже для современных УЗИ-аппаратов.
Врач-узист обращает внимание не только на размер, но и на форму плодного яйца. Идеальная форма — правильный овал или круг. Если плодное яйцо имеет неправильную форму или его контуры нечеткие, это может быть поводом для беспокойства и требует дополнительного наблюдения.
Расположение плодного яйца также имеет значение. В норме оно должно находиться в верхней части полости матки, примерно посередине. Если плодное яйцо расположено слишком низко или сбоку, это может указывать на проблемы с имплантацией.
Что дальше? Развитие после 5 мм
Обнаружение 5 мм плодного яйца — это только начало удивительного путешествия. В ближайшие недели темпы развития будут поистине ошеломляющими. Вот что произойдет в ближайшее время:
- Плодное яйцо будет быстро увеличиваться в размерах, примерно на 1 мм в день.
- Через неделю после обнаружения 5 мм плодного яйца уже можно будет увидеть эмбрион и, возможно, даже услышать его сердцебиение!
- К 6-7 неделе беременности у эмбриона начнут формироваться зачатки глаз, ушей и даже пальчиков.
- К 8 неделе эмбрион уже будет иметь все основные органы и системы, хотя они еще не полностью развиты.
Возможные сложности и как их распознать
К сожалению, не всегда все идет по плану. Иногда даже при обнаружении 5 мм плодного яйца могут возникнуть проблемы. Вот на что стоит обратить внимание:
- Анэмбриония: ситуация, когда плодное яйцо есть, но эмбрион внутри него не развивается. В этом случае при повторном УЗИ через 1-2 недели внутри увеличившегося плодного яйца не будет видно эмбриона.
- Замершая беременность: эмбрион есть, но его развитие остановилось. При этом размеры эмбриона не соответствуют сроку беременности, а сердцебиение отсутствует.
- Внематочная беременность: плодное яйцо имплантировалось не в матке, а например, в маточной трубе. Это опасное состояние, требующее немедленного медицинского вмешательства.
Важно помнить, что наличие 5 мм плодного яйца — это хороший знак, но не гарантия успешного развития беременности. Поэтому так важно регулярное наблюдение у врача и выполнение всех его рекомендаций.
Эмоциональный аспект: первая встреча с малышом
Обнаружение 5 мм плодного яйца на УЗИ — это не просто медицинский факт. Для многих будущих родителей это первая «встреча» со своим малышом, пусть пока и виртуальная. Этот момент часто вызывает бурю эмоций: радость, волнение, а иногда и страх перед неизвестностью.
Многие женщины отмечают, что именно с этого момента они начинают ощущать себя будущими мамами. Появляется желание заботиться о себе, менять привычки, готовиться к предстоящим изменениям в жизни. И хотя 5 мм плодное яйцо еще очень мало, оно уже способно перевернуть весь мир будущих родителей!
Таким образом, путь от оплодотворения до формирования 5 мм плодного яйца — это удивительное путешествие, полное чудес и открытий. Каждый день приносит новые изменения, приближая момент, когда крошечная клетка превратится в нового человека. И хотя впереди еще долгий путь, обнаружение 5 мм плодного яйца — это первый важный шаг, первое реальное доказательство того, что новая жизнь уже начала свой путь.
Симптомы на ранних сроках: Что чувствует женщина при 5 мм плодном яйце
Ранняя беременность — это время, когда в организме женщины происходят колоссальные изменения, хотя внешне они могут быть совершенно незаметны. Когда плодное яйцо достигает размера 5 мм, что обычно происходит на 4-5 неделе беременности, многие женщины уже начинают замечать первые признаки своего «интересного положения». Однако стоит отметить, что симптомы могут сильно варьироваться от женщины к женщине, и даже от беременности к беременности у одной и той же женщины.
Итак, что же может чувствовать будущая мама, когда внутри нее формируется крошечное 5 мм плодное яйцо? Давайте разберемся в этом вопросе, опираясь на опыт множества женщин и научные данные.
Отсутствие менструации: Первый звоночек
Самый очевидный и часто первый замеченный симптом — это задержка менструации. Когда плодное яйцо достигает размера 5 мм, обычно уже прошло несколько дней с момента ожидаемого начала месячных. Однако не все женщины сразу обращают на это внимание, особенно если у них нерегулярный цикл. Кроме того, некоторые могут спутать имплантационное кровотечение с началом менструации. Поэтому важно быть внимательной к своему телу и вести календарь менструального цикла.
Грудь: Источник первых ощущений
Многие женщины отмечают, что одним из первых признаков беременности становятся изменения в груди. Что же происходит? Грудь может стать более чувствительной, даже болезненной при прикосновении. Некоторые описывают это ощущение как «покалывание» или «тяжесть». Кроме того, вы можете заметить, что ареолы (область вокруг сосков) стали темнее и больше. Эти изменения связаны с гормональной перестройкой организма, которая начинается сразу после имплантации плодного яйца.
Утренняя тошнота: Миф или реальность?
Вопреки распространенному мнению, не все женщины испытывают утреннюю тошноту на ранних сроках беременности. Когда плодное яйцо достигает размера 5 мм, некоторые будущие мамы уже могут чувствовать легкое недомогание, особенно по утрам. Однако многие в этот период еще не испытывают никакого дискомфорта. Интересно, что утренняя тошнота может проявляться не только утром — некоторые женщины отмечают, что чувствуют дискомфорт в течение всего дня или даже вечером.

Усталость: Неожиданный спутник
Многие женщины отмечают, что уже на самых ранних сроках беременности, когда плодное яйцо еще совсем крошечное — всего 5 мм, они начинают чувствовать необычайную усталость. Это может проявляться как постоянное желание прилечь, трудности с концентрацией внимания или просто ощущение, что «батарейка села». Такая усталость связана с гормональными изменениями и тем, что организм начинает перестраиваться для поддержки новой жизни. Не стоит с этим бороться — если чувствуете усталость, постарайтесь больше отдыхать.
Перепады настроения: Эмоциональные американские горки
Гормональные изменения, происходящие в организме женщины при формировании 5 мм плодного яйца, могут вызвать настоящие эмоциональные «американские горки». Вы можете заметить, что стали более чувствительны, легко расстраиваетесь или, наоборот, испытываете необъяснимую эйфорию. Некоторые женщины отмечают, что могут заплакать из-за трогательной рекламы или расстроиться из-за мелочи, которая раньше их не беспокоила. Это нормально! Ваш организм адаптируется к новому состоянию, и эмоции могут быть нестабильными.
Изменения в пищевых предпочтениях: Странные желания
Хотя классические «беременные причуды» обычно появляются позже, некоторые женщины отмечают изменения в пищевых предпочтениях уже на ранних сроках, когда плодное яйцо достигает размера 5 мм. Это может проявляться как внезапное отвращение к ранее любимым продуктам или, наоборот, сильное желание съесть что-то необычное. Например, женщина, которая никогда не любила острую пищу, может вдруг обнаружить, что ей хочется съесть острый перец. Или наоборот, кофеманка может заметить, что запах кофе вызывает у нее тошноту.
Частое мочеиспускание: Ранний признак
Многие женщины удивляются, узнав, что частые походы в туалет могут начаться уже на очень ранних сроках беременности, когда плодное яйцо еще совсем маленькое — всего 5 мм. Это связано с тем, что увеличивающаяся матка начинает давить на мочевой пузырь, а также с изменениями в работе почек под влиянием гормонов беременности. Если вы заметили, что стали чаще бегать в туалет, особенно ночью, это может быть одним из первых признаков беременности.
Головокружение и обмороки: Неожиданные спутники
Некоторые женщины отмечают, что уже на ранних сроках беременности, когда плодное яйцо достигает размера 5 мм, они начинают испытывать легкое головокружение или даже склонность к обморокам. Это связано с расширением кровеносных сосудов под действием гормонов беременности, что может привести к временному снижению артериального давления. Если вы почувствовали головокружение, лучше присесть или прилечь, чтобы избежать падения.
Повышенная чувствительность к запахам: Обостренное обоняние
Удивительно, но многие женщины отмечают, что их обоняние становится сверхчувствительным уже на ранних сроках беременности. Когда плодное яйцо достигает размера 5 мм, некоторые будущие мамы могут обнаружить, что запахи, которые раньше их не беспокоили, вдруг стали невыносимыми. Или наоборот, появляется сильная тяга к определенным ароматам. Это может быть связано с повышением уровня эстрогена в организме.
Изменения кожи: Первые признаки «цветения»
Хотя классическое «сияние беременности» обычно появляется позже, некоторые женщины отмечают изменения кожи уже на ранних сроках, когда плодное яйцо достигает размера 5 мм. У кого-то кожа становится более склонной к высыпаниям, у других, наоборот, проблемы с кожей исчезают. Некоторые женщины замечают появление пигментных пятен или потемнение кожи в определенных местах (например, вокруг сосков или на линии живота).
Боли и спазмы: Нормально ли это?
Многие женщины испытывают легкие боли или спазмы в нижней части живота на ранних сроках беременности, когда плодное яйцо достигает размера 5 мм. Эти ощущения часто похожи на менструальные боли и связаны с растяжением матки и имплантацией плодного яйца. Однако если боль сильная или сопровождается кровотечением, необходимо немедленно обратиться к врачу.
Отсутствие симптомов: Повод для беспокойства?
Важно помнить, что отсутствие каких-либо симптомов на ранних сроках беременности, когда плодное яйцо достигает размера 5 мм, — это тоже вариант нормы. Некоторые женщины не испытывают никаких необычных ощущений вплоть до нескольких месяцев беременности. Это не означает, что с беременностью что-то не так. Каждая женщина и каждая беременность уникальны, и отсутствие симптомов не является поводом для беспокойства.
В заключение стоит отметить, что симптомы на ранних сроках беременности, когда плодное яйцо достигает размера 5 мм, могут быть очень разнообразными и индивидуальными. Некоторые женщины могут испытывать все вышеперечисленные симптомы, другие — только некоторые из них, а третьи могут не чувствовать ничего необычного. В любом случае, если вы подозреваете, что беременны, лучше всего сделать тест и обратиться к врачу для подтверждения беременности и начала наблюдения.
Диагностика и наблюдение: Важность регулярных осмотров при маленьком плодном яйце
Что такое это загадочное 5 мм плодное яйцо, о котором так часто говорят врачи? Давайте разберемся! Представьте себе крошечный мешочек, размером с горошину, который появляется в матке на ранних сроках беременности. Вот это и есть плодное яйцо — колыбель новой жизни, которая только начинает формироваться.
Но почему же такое внимание уделяется именно размеру в 5 мм? Дело в том, что это своего рода критическая точка. При нормальном развитии беременности плодное яйцо должно быть больше. Если оно остается таким маленьким, это может быть сигналом, что что-то идет не так. Но не паникуйте раньше времени! Иногда это просто означает, что срок беременности меньше, чем предполагалось изначально.
Как же выявляют это миниатюрное плодное яйцо? Главный помощник здесь — УЗИ. Этот волшебный аппарат позволяет заглянуть внутрь матки и увидеть то, что скрыто от глаз. При этом исследовании врач может измерить размер плодного яйца с точностью до миллиметра. Удивительно, правда? Современные технологии позволяют нам наблюдать за самым началом жизни!
Но не стоит думать, что 5 мм плодное яйцо — это приговор. Каждая беременность уникальна, и развитие может идти по-разному. Иногда плодное яйцо просто не успело подрасти к моменту обследования. В таких случаях врачи обычно назначают повторное УЗИ через неделю-две. За это время ситуация может кардинально измениться.
А какие еще признаки могут указывать на раннюю беременность, кроме маленького плодного яйца? Ну, во-первых, это задержка менструации. Многие женщины именно по этому признаку начинают подозревать, что они в положении. Во-вторых, это тест на беременность, который показывает две заветные полоски. Но бывает, что тест положительный, а на УЗИ видно только крошечное 5 мм плодное яйцо. Не спешите расстраиваться! Это может быть просто очень ранний срок.
Еще один интересный момент — это ощущения самой женщины. Некоторые будущие мамы утверждают, что чувствуют изменения в своем теле буквально с первых дней после зачатия. Кто-то жалуется на тошноту и головокружение, кто-то отмечает повышенную чувствительность к запахам. А кто-то и вовсе не замечает никаких изменений, пока не увидит на мониторе УЗИ это самое 5 мм плодное яйцо!
Но вернемся к нашему маленькому герою. Что же происходит внутри этого крошечного мешочка? На самом деле, там кипит жизнь! Уже на этом этапе начинают формироваться важнейшие органы и системы будущего малыша. Представляете, какая колоссальная работа происходит в таком микроскопическом пространстве?
А знаете ли вы, что размер плодного яйца может многое рассказать о сроке беременности? Например, 5 мм плодное яйцо обычно соответствует 4-5 неделям беременности. Это тот самый момент, когда большинство женщин только-только узнают о своем интересном положении. Удивительно, как природа все точно рассчитала!
Но что делать, если врач обнаружил 5 мм плодное яйцо, а по расчетам срок беременности должен быть больше? В такой ситуации важно сохранять спокойствие и следовать рекомендациям специалиста. Обычно назначают повторное УЗИ через некоторое время, чтобы оценить динамику роста. Кроме того, могут быть назначены анализы на уровень гормонов, которые помогут лучше оценить ситуацию.
Интересно, что иногда маленькое плодное яйцо может быть связано с многоплодной беременностью. Бывает, что на раннем сроке врач видит два маленьких плодных яйца вместо одного большого. Представляете, какой сюрприз ждет будущих родителей? Двойное счастье!
А как же быть с волнением и тревогой, которые неизбежно возникают при обнаружении маленького плодного яйца? Психологи советуют сосредоточиться на позитивных моментах. Например, сам факт наличия плодного яйца уже говорит о том, что беременность наступила и развивается. Это уже повод для радости!
Кстати, а вы знали, что размер плодного яйца может зависеть от многих факторов? Например, от индивидуальных особенностей организма женщины, от ее образа жизни и питания. Поэтому так важно еще до беременности заботиться о своем здоровье и готовить организм к этому важному этапу.
Но давайте вернемся к нашему 5 мм плодному яйцу. Что же происходит дальше, если все идет хорошо? Буквально через неделю-две оно может вырасти до 10-15 мм. А еще через некоторое время внутри него можно будет разглядеть крошечный эмбрион. Удивительно, как быстро развивается новая жизнь!
В заключение хочется сказать: если вы столкнулись с ситуацией, когда на УЗИ обнаружили 5 мм плодное яйцо, не паникуйте. Это может быть просто началом прекрасного путешествия под названием «беременность». Главное — внимательно следить за своим здоровьем, выполнять рекомендации врача и верить в лучшее. Ведь каждая новая жизнь — это маленькое чудо, даже если она начинается с такого крошечного плодного яйца!
Возможные риски: Когда 5 мм плодное яйцо может вызывать беспокойство
Когда речь заходит о 5 мм плодном яйце, многие будущие мамы начинают нервничать. И неспроста! Этот крошечный объект, едва различимый на УЗИ, может стать причиной серьезных волнений. Но давайте разберемся, когда действительно стоит бить тревогу, а когда можно выдохнуть с облегчением.
Прежде всего, важно понимать: размер плодного яйца — это не приговор, а лишь один из показателей. Иногда 5 мм плодное яйцо — это просто признак очень ранней беременности. Но бывают ситуации, когда такой размер на определенном сроке может указывать на проблемы. Как же отличить одно от другого?
Представьте себе, что плодное яйцо — это маленький космический корабль, который только начал свое путешествие. Если он слишком долго остается маленьким, это может означать, что у него не хватает топлива для полета. В нашем случае «топливо» — это правильный гормональный фон и благоприятные условия в матке.
Одна из главных причин для беспокойства — это несоответствие размера плодного яйца предполагаемому сроку беременности. Если по расчетам беременность уже 7-8 недель, а плодное яйцо все еще 5 мм, это может быть признаком замершей беременности. Но не спешите паниковать! Иногда это просто означает, что овуляция произошла позже, чем предполагалось, и беременность на самом деле «моложе».
Другой повод для волнения — отсутствие роста плодного яйца при повторных УЗИ. Если через неделю-две после первого обследования размер не изменился, это уже серьезный сигнал. В норме плодное яйцо должно расти примерно на 1 мм в день. Если этого не происходит, врачи начинают подозревать анэмбрионию — ситуацию, когда плодное яйцо есть, а эмбрион внутри не развивается.
А что насчет формы? Оказывается, не только размер, но и форма плодного яйца может многое рассказать. Идеальное плодное яйцо должно быть круглым или слегка овальным. Если же оно имеет неправильную форму или нечеткие контуры, это может указывать на проблемы с имплантацией или развитием.
Интересный факт: иногда маленькое плодное яйцо может быть признаком многоплодной беременности на самом раннем сроке. Представьте, врач видит два маленьких плодных яйца вместо одного большого. Вот это поворот, правда? Хотя, конечно, такая ситуация встречается реже, чем другие причины маленького размера плодного яйца.
Еще один момент, который может вызвать беспокойство — это отсутствие желточного мешка внутри 5 мм плодного яйца. Обычно желточный мешок должен быть виден, когда плодное яйцо достигает размера 8-10 мм. Если его нет, это может указывать на нарушения развития. Но опять же, не стоит делать поспешных выводов — иногда желточный мешок просто не виден из-за особенностей расположения плодного яйца.
А как насчет гормонального фона? Уровень ХГЧ (хорионического гонадотропина человека) в крови может многое рассказать о состоянии беременности. Если плодное яйцо маленькое, а уровень ХГЧ низкий или не растет должным образом, это повод для дополнительного обследования. С другой стороны, нормальный рост ХГЧ при маленьком плодном яйце может говорить о том, что беременность просто на более раннем сроке, чем предполагалось изначально.
Кстати, о сроках. Знаете ли вы, что определение точного срока беременности по последней менструации не всегда точно? Иногда овуляция происходит позже или раньше, чем ожидалось. Поэтому 5 мм плодное яйцо, которое кажется маленьким для предполагаемого срока, может на самом деле соответствовать реальному возрасту беременности.
Еще один фактор, который нужно учитывать — это качество УЗИ-аппарата и опыт врача. Иногда маленький размер плодного яйца может быть результатом погрешности измерения или использования устаревшего оборудования. Вот почему так важно проходить обследование в проверенных клиниках с современной аппаратурой.
А что делать, если 5 мм плодное яйцо действительно вызывает обоснованное беспокойство? Врачи обычно рекомендуют повторное УЗИ через 7-10 дней. За это время нормально развивающееся плодное яйцо должно значительно увеличиться в размерах. Если рост не наблюдается, могут быть назначены дополнительные исследования, включая анализ на ХГЧ и прогестерон.
Интересно, что иногда маленькое плодное яйцо может быть связано с генетическими аномалиями. Исследования показывают, что в случаях, когда размер плодного яйца значительно меньше нормы для данного срока, риск хромосомных нарушений выше. Однако это не значит, что каждое маленькое плодное яйцо — это приговор. Многие беременности с изначально маленьким плодным яйцом заканчиваются рождением совершенно здоровых малышей.
А как же психологический аспект? Стресс от неопределенности может негативно влиять на течение беременности. Поэтому так важно найти баланс между бдительностью и спокойствием. Некоторые врачи рекомендуют техники релаксации и медитации для снижения уровня тревоги. Ведь, в конце концов, стресс не поможет плодному яйцу вырасти быстрее, правда?
Еще один интересный момент — влияние образа жизни на развитие плодного яйца. Хотя прямой связи между размером плодного яйца и образом жизни женщины не установлено, общее состояние здоровья может играть роль. Правильное питание, отказ от вредных привычек, умеренные физические нагрузки — все это создает благоприятный фон для развития беременности.
А знаете ли вы, что иногда маленькое плодное яйцо может быть признаком внематочной беременности? Хотя это редкий случай, но его нельзя исключать. Если 5 мм плодное яйцо не визуализируется в матке, но есть другие признаки беременности, врачи обязательно проверят маточные трубы.
В заключение хочется сказать: если вы столкнулись с ситуацией, когда обнаружено 5 мм плодное яйцо, и это вызывает у вас беспокойство, помните — вы не одни. Многие женщины проходят через это. Главное — не паниковать раньше времени, внимательно следовать рекомендациям врача и помнить, что каждая беременность уникальна. То, что кажется проблемой сегодня, завтра может обернуться радостной новостью о здоровом развитии вашего будущего малыша.
Питание и образ жизни: Рекомендации для здорового развития эмбриона
Когда УЗИ показывает 5 мм плодное яйцо, будущие мамы часто задаются вопросом: «А что я могу сделать, чтобы помочь моему крохе расти здоровым?». И знаете что? Это отличный вопрос! Ведь даже на таком раннем этапе, когда эмбрион едва различим, наши действия могут сыграть важную роль в его развитии.
Начнем с самого очевидного — питания. Помните, как в детстве нам говорили: «Ты то, что ты ешь»? Так вот, для беременных это вдвойне верно! Ваш рацион — это строительный материал для растущего организма. Но что же конкретно нужно есть, когда у вас обнаружили 5 мм плодное яйцо?
Прежде всего, забудьте о диетах! Сейчас не время считать калории. Ваш организм нуждается в питательных веществах, чтобы поддержать развитие эмбриона. Фолиевая кислота, железо, кальций — вот ваши лучшие друзья в этот период. Где их найти? В зеленых листовых овощах, бобовых, цитрусовых. А как насчет белка? Он необходим для формирования тканей плода. Нежирное мясо, рыба, яйца — отличный выбор.
Но питание — это не только «что», но и «как». Маленькие порции, частые приемы пищи — вот золотое правило. Представьте, что вы кормите не только себя, но и крошечный «реактор» внутри, которому нужно постоянное топливо. 5 мм плодное яйцо может казаться малышом, но его аппетит уже вполне взрослый!
А как насчет витаминов? Многие врачи рекомендуют принимать пренатальные витамины еще до зачатия. Если вы еще не начали, самое время начать сейчас. Но внимание! Не стоит заниматься самолечением. Выбор витаминных комплексов должен быть согласован с вашим врачом.
Теперь поговорим о жидкости. Вода — источник жизни, и это особенно верно во время беременности. Достаточное потребление воды помогает формированию амниотической жидкости, которая окружает и защищает ваше 5 мм плодное яйцо. Сколько пить? Минимум 8 стаканов в день, а лучше больше.
А что насчет кофе? Многие будущие мамы паникуют, узнав о беременности, и резко отказываются от любимого напитка. Но не спешите! Умеренное потребление кофе (до 200 мг кофеина в день) считается безопасным. Это примерно одна чашка фильтр-кофе или два эспрессо. Но помните, кофеин содержится не только в кофе, но и в чае, шоколаде, некоторых газированных напитках.
Теперь давайте поговорим о физической активности. Многие женщины, узнав о 5 мм плодном яйце, боятся лишний раз пошевелиться. Но это ошибка! Умеренные физические нагрузки не только безопасны, но и полезны для развития эмбриона. Они улучшают кровообращение, помогают бороться со стрессом, укрепляют мышцы. Йога для беременных, плавание, пешие прогулки — отличный выбор. Но помните о золотой середине — не стоит внезапно становиться марафонцем, если раньше вы вели сидячий образ жизни.
Кстати, о сидячем образе жизни. Если ваша работа предполагает долгое сидение за компьютером, обязательно делайте перерывы. Встаньте, пройдитесь, сделайте легкую разминку. Это поможет улучшить кровообращение, что крайне важно для развития вашего 5 мм плодного яйца.
А как насчет сна? О, это отдельная история! Казалось бы, какая связь между сном и развитием эмбриона? Самая прямая! Во время сна ваш организм восстанавливается, производит важные гормоны, необходимые для роста плода. Старайтесь спать не менее 8 часов в сутки. И да, дневной сон — это не лень, а забота о малыше!
Теперь поговорим о стрессе. Знаете ли вы, что высокий уровень стресса может негативно влиять на развитие эмбриона? Да, даже когда плодное яйцо всего 5 мм! Поэтому так важно научиться расслабляться. Медитация, дыхательные упражнения, приятная музыка — выберите то, что подходит именно вам.
А как насчет вредных привычек? Думаю, вы и сами знаете ответ. Алкоголь, курение, наркотики — это абсолютное табу во время беременности. Даже если вы узнали о беременности недавно и плодное яйцо еще совсем маленькое, немедленно откажитесь от этих привычек. Ваш малыш скажет вам спасибо!
Интересный факт: некоторые исследования показывают, что музыка может положительно влиять на развитие эмбриона. Хотя ваше 5 мм плодное яйцо еще не имеет ушей, вибрации могут восприниматься на клеточном уровне. Так что включайте любимую классику — пусть малыш приобщается к прекрасному с самых первых дней!
А как насчет секса? Многие пары боятся интимной близости, узнав о беременности. Но если беременность протекает нормально, и нет противопоказаний от врача, то секс безопасен даже на ранних сроках. Более того, он может помочь снять напряжение и улучшить эмоциональное состояние.
Теперь поговорим об окружающей среде. Пусть ваше 5 мм плодное яйцо еще крошечное, но оно уже нуждается в защите. Старайтесь избегать контакта с вредными химическими веществами, не дышите табачным дымом, будьте осторожны с бытовой химией. Может быть, стоит задуматься об экологичных средствах для уборки?
Еще один важный момент — эмоциональный настрой. Позитивное мышление — не просто красивые слова. Исследования показывают, что эмоциональное состояние матери влияет на развитие плода. Так что улыбайтесь чаще, общайтесь с приятными людьми, занимайтесь тем, что приносит вам радость.
И наконец, не забывайте о регулярных визитах к врачу. Даже если ваше плодное яйцо всего 5 мм, важно следить за его развитием. Регулярные осмотры помогут вовремя заметить любые отклонения и принять необходимые меры.
В заключение хочется сказать: беременность — это удивительное время. Даже когда плодное яйцо еще совсем крошечное, всего 5 мм, вы уже можете многое сделать для здоровья будущего малыша. Правильное питание, умеренная физическая активность, отказ от вредных привычек, позитивный настрой — все это закладывает фундамент здоровья вашего ребенка. Помните, каждый день вы делаете важную работу — растите новую жизнь. И пусть эта работа приносит вам радость и удовлетворение!
Эмоциональное состояние: Как справиться с волнением на раннем сроке беременности
Узнав о том, что УЗИ показало 5 мм плодное яйцо, многие женщины испытывают целую гамму эмоций. Радость смешивается с тревогой, восторг — с неуверенностью. И это абсолютно нормально! Ведь внутри вас зарождается новая жизнь, а это, согласитесь, не шутки. Как же справиться с этим эмоциональным вихрем и не дать тревоге взять верх?
Прежде всего, важно понимать, что ваши чувства — это не просто каприз гормонов. Это естественная реакция на серьезные изменения в вашей жизни. Представьте, что вы начинаете новый грандиозный проект. Разве не будете вы волноваться о его успехе? А беременность — это самый важный проект в жизни женщины!
Одна из главных причин беспокойства на раннем сроке — это страх потери беременности. Когда плодное яйцо всего 5 мм, кажется, что оно такое хрупкое и уязвимое. Но помните: природа мудра. Она создала множество механизмов для защиты развивающейся жизни. Ваш организм знает, что делать, даже если вы сами этого не осознаете.
Как же бороться с тревогой? Один из эффективных методов — это глубокое дыхание. Звучит просто, но работает потрясающе! Когда вы чувствуете, что волнение нарастает, сделайте несколько глубоких вдохов и выдохов. Представьте, как с каждым вдохом вы наполняетесь спокойствием, а с каждым выдохом отпускаете тревогу. Это не только успокаивает нервы, но и улучшает кровоснабжение, что полезно для вашего 5 мм плодного яйца.
Другой способ справиться с эмоциями — это разговор. Не держите свои переживания в себе! Поделитесь ими с партнером, подругой или мамой. Иногда просто проговаривание своих страхов вслух помогает увидеть ситуацию в другом свете. А если рядом нет никого, кому вы могли бы довериться? Заведите дневник беременности. Записывайте туда свои мысли, чувства, ожидания. Это не только поможет выплеснуть эмоции, но и станет прекрасным воспоминанием в будущем.
Знаете, что еще помогает справиться с тревогой? Информация! Когда мы чего-то не знаем, мы начинаем придумывать самые страшные сценарии. Поэтому читайте, изучайте, спрашивайте у врача. Узнайте, что происходит с вашим телом, как развивается ваше 5 мм плодное яйцо. Но будьте осторожны: интернет полон страшилок и непроверенной информации. Выбирайте надежные источники и не зацикливайтесь на негативных историях.
А как насчет позитивной визуализации? Это мощный инструмент для борьбы со стрессом. Закройте глаза и представьте свое плодное яйцо. Да, оно всего 5 мм, но оно растет и развивается. Представьте, как оно окружено любовью и заботой. Как внутри него формируется новая жизнь. Такие «медитации» не только успокаивают, но и помогают установить эмоциональную связь с будущим малышом.
Еще один важный момент — это физическая активность. Нет, речь не о марафонах и тяжелой атлетике. Легкая йога, пешие прогулки, плавание — все это отлично снимает стресс и улучшает настроение. Когда мы двигаемся, наш мозг вырабатывает эндорфины — гормоны счастья. А счастливая мама — это счастливое 5 мм плодное яйцо!
Кстати, о счастье. Не забывайте о маленьких радостях жизни! Беременность — это не болезнь, и вы не обязаны сидеть дома и постоянно думать о своем состоянии. Ходите в кино, встречайтесь с друзьями, занимайтесь хобби. Все это поможет отвлечься от тревожных мыслей и насладиться этим особенным периодом вашей жизни.
А что делать, если тревога становится слишком сильной? Не стесняйтесь обратиться за профессиональной помощью. Многие женские консультации предлагают консультации психолога для будущих мам. Это не признак слабости, а наоборот, проявление заботы о себе и своем малыше.
Интересный факт: некоторые исследования показывают, что эмоциональное состояние матери может влиять на развитие плода. Поэтому, борясь со стрессом, вы не только улучшаете свое самочувствие, но и создаете благоприятную среду для роста вашего 5 мм плодного яйца.
Еще один способ справиться с тревогой — это планирование. Когда мы чувствуем, что контролируем ситуацию, нам становится спокойнее. Начните готовиться к будущему материнству уже сейчас. Составьте список вещей, которые нужно купить для малыша, подумайте об обустройстве детской комнаты. Да, до родов еще далеко, но такое приятное планирование поможет переключить фокус с тревоги на позитивные мысли о будущем.
А как насчет творчества? Многие женщины отмечают, что во время беременности у них просыпаются скрытые таланты. Может быть, вы всегда хотели научиться рисовать или писать стихи? Сейчас самое время попробовать! Творческий процесс не только отвлекает от тревожных мыслей, но и помогает выразить свои эмоции в позитивном ключе.
Не забывайте и о поддержке партнера. Часто мужчины чувствуют себя немного «за бортом» на ранних сроках беременности, особенно когда плодное яйцо еще такое маленькое — всего 5 мм. Расскажите ему о своих чувствах, поделитесь информацией о развитии малыша. Это поможет ему почувствовать себя более вовлеченным, а вам — получить необходимую поддержку.
И последнее, но не менее важное: будьте добры к себе. Беременность — это время больших изменений, и вполне нормально чувствовать себя немного неуверенно. Не ругайте себя за тревожные мысли или смену настроения. Примите свои чувства, какими бы они ни были. Помните: вы делаете великое дело — растите новую жизнь. И пусть ваше плодное яйцо сейчас всего 5 мм, но с каждым днем оно растет, как и ваша сила и уверенность в себе как в будущей маме.